Сиволоб А.В., Рушковський С.Р., Кир’яченко С.С. та ін. Генетика. Підручник
Подождите немного. Документ загружается.

Розділ 5. Генетика бактерій, вірусів і одноклітинних еукаріотів
161
штаму не розмножуються на середовищі без триптофану, а іншого –
потребують гістидин, то на відповідному середовищі можна відібрати
"гібридні" диплоїдні колонії.
2n
nn
n n
n
n
n
n
a α
2n
2n
2n
2n
n
n
Д
иплоїдна клітина,
що брунькується
Аск зі спорами
Гаплоїдна клітина,
що брунькується
Рис. 5.10. Життєвий цикл аскоміцета Saccharomyces cerevisiae
Якщо ізолювати окрему аскоспору, то її потомство, яке виникне за
рахунок брунькування, має складатися з клітин одного типу. Проте та-
кий штам не буде насправді моноспоровим, якщо в нього присутній
ген НО, який забезпечує перемикання типів клітин: клітина а утворює
бруньку типу α (або навпаки). Звичайно, у цьому випадку клітини різно-
го типу, що накопичуються, будуть спарюватися з утворенням диплоїд-
них гетерозигот, які врешті-решт і будуть переважати в культурі.
Перемикання типів клітин у S. cerevisiae є прикладом програмо-
ваної геномної перебудови. Локус МАТ розташований у правому плечі
третьої хромосоми: елемент послідовності Yа або Yα, який визначає
алельну форму локусу, фланкований кількома іншими елементами
з обох боків (рис. 5.11, де зображено конфігурацію, що відповідає
алелю МАТα). Поблизу від лівої та правої теломер є дві так звані касети,
що являють собою алелі α та а відповідно. Але касети (позначаються
як HMLα та HMLа) знаходяться у субтеломерних гетерохроматинових
зонах, і тому не експресуються – вони недоступні для системи транс-
крипції внаслідок додаткової компактизації хроматину (див. розділ 6).
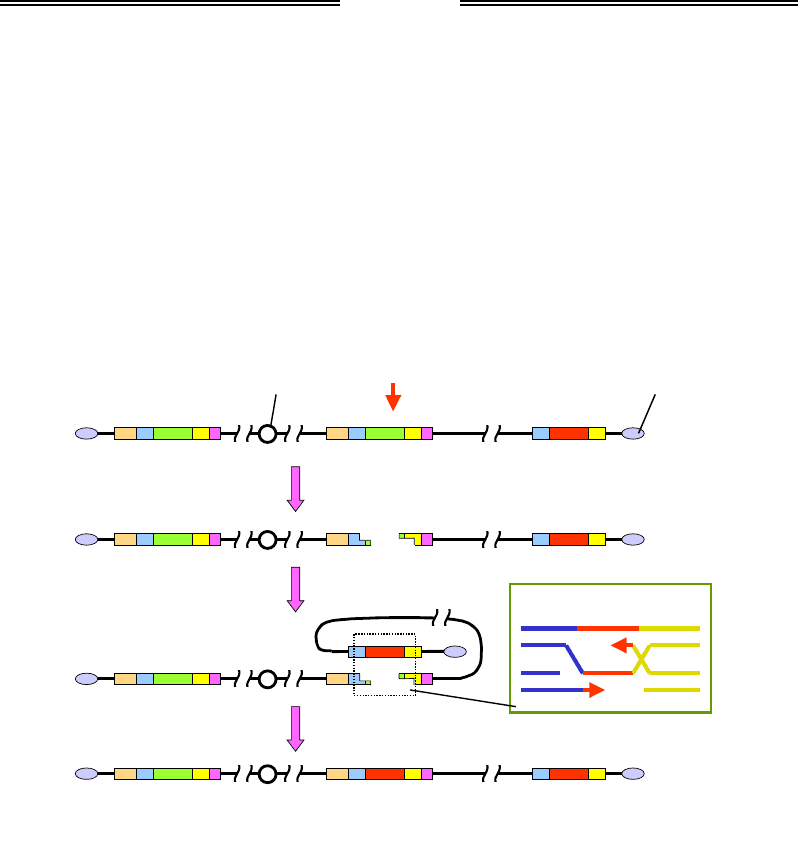
Генетика
162
При перемиканні типів клітин спрацьовує ген НО, який кодує спе-
цифічну нуклеазу. Нуклеаза НО розрізає локус МАТ у зоні елемента Yα
(або Yа). Наступні події еквівалентні схемі гомологічної рекомбінації
(порівн. рис. 5.11 і 1.25): дволанцюговий розріз розширюється
з утворенням двох 3'-кінцевих одноланцюгових хвостів, у хромосомі
утворюється петля й гомологічна касета HMLа підводиться до зони
розрізу, відбувається репараційний синтез ДНК на ланцюгах ДНК ка-
сети. Результатом є конверсія гена: заміна локусу МАТα на локус МАТа.
Обидві касети при цьому залишаються в незмінному вигляді, і в май-
бутніх поколіннях ліва касета може бути використана для зворотного
перемикання локусу МАТа на МАТα.
WX
Yα
Z1
Z2
WX
Yα
Z1
Z2
X
Ya
Z1
HMLα
HMLaMATα
MATa
X
Ya
Z1
Центромера Теломера
HO
Рис. 5.11. Схема перемикання типу спарювання у S. cerevisiae.
Якщо в центрі міститься локус МАТа, для конверсії використовується касета HMLα
Результатом синтезу ДНК, що зображено на вставці рис. 5.11, буде
утворення двох структур Холідея (порівн. рис. 1.26). Як пояснювалось
у розділі 1, є два рівноймовірні шляхи розділення цих структур: із кро-
синговером між двома дуплексами та без (див. також рис. 1.28).
У випадку, зображеному на рис. 5.11, кросинговер між двома дуплек-
сами, що належать одній хромосомі, зумовить делецію – вирізання ді-
лянки між локусом МАТ і касетою HMLа. Отже, у цьому разі спрямо-

Розділ 5. Генетика бактерій, вірусів і одноклітинних еукаріотів
163
вано реалізуються лише такі варіанти розділення, які не приводять до
кросинговеру та, відповідно, хромосомних аберацій.
Слід зауважити, що й при міжхроматидній гомологічній рекомбі-
нації не завжди кросинговер і конверсія без кросинговеру є рівно-
ймовірними подіями. Так, при рекомбінації під час мітозу диплоїдних
клітин дріжджів кросинговер відбувається лише в 10 % рекомбіна-
ційних подій. Таким чином, принаймні в деяких випадках, процеси
гомологічної рекомбінації можуть здійснюватися саме з метою конвер-
сії геномної ділянки, яка б не супроводжувалась кросинговером.
Касетний механізм перемикання активності генів є досить поши-
реним: аналогічні процеси описано для інших видів аскоміцетів, три-
паносом і деяких бактерій. Крім того, такий механізм напевно реалі-
зується при внутрішньохромосомній гомологічній рекомбінації на ге-
нах, що тандемно повторюються (за принципом, який ілюструє
рис. 5.11, і без кросинговеру), з метою підтримувати ідентичність тан-
демних копій: мутація в одній з таких копій з високою імовірністю
буде елімінована за рахунок використання іншої з багатьох нормаль-
них гомологічних ділянок як матриці.
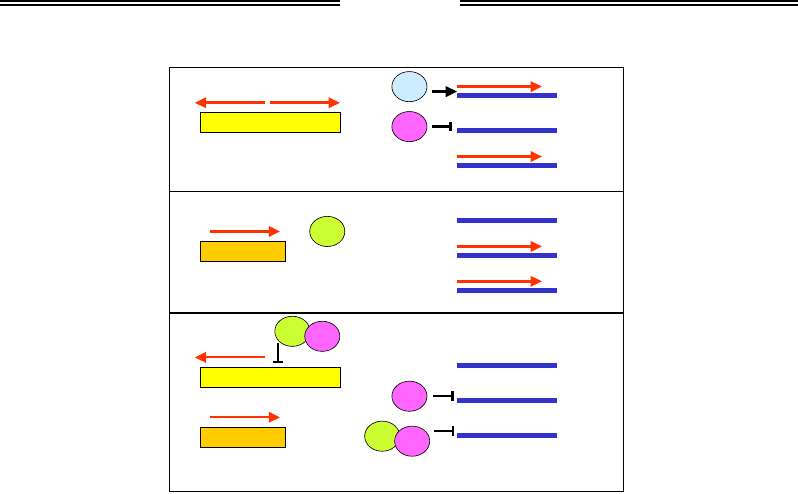
Повернемося до перемикання типів клітин у S. сerevisiae, яке є та-
кож добре вивченим прикладом взаємодії генів на рівні регуляції транс-
крипції. Приблизно по центру локусу МАТα розташована операторна ді-
лянка, що активує два промотори, з яких відбувається транскрипція
у двох протилежних напрямках. Продуктами цих генів є два білки α1 та
α2 – транскрипційні фактори. Перший із них активує транскрипцію
групи генів, які визначають специфічні ознаки клітин α-типу, другий –
є репресором для групи а-специфічних генів (рис. 5.12). При цьому в га-
плоїдних клітинах обох типів експресується група генів, специфічних
для гаплоїдних клітин узагалі – за рахунок активації іншим фактором
транскрипції. У клітинах а-типу з локусу МАТа експресується білок а1,
який сам по собі не впливає на транскрипцію зазначених груп генів, але
відсутність білків α1 і α2 зумовлює активацію а-специфічних (за відсут-
ності репресора) і вимикання α-специфічних (за відсутності активатора)
генів (рис. 5.12). У гетерозиготних за обома МАТ-локусами диплоїдних
клітинах білок а1 утворює комплекс із α2, який ефективно блокує
транскрипцію α1 (у результаті α-специфічні гени знову вимкнено), а та-
кож транскрипцію генів, специфічних для гаплоїдних клітин (рис. 5.12).
Крім того, комплекс а1-α2 є репресором гена НО, який не здатен
ініціювати процес рекомбінації, зображений на рис. 5.11, у диплоїд-
них клітинах. Активація цього гена в гаплоїдних клітинах потребує
набору певних активаторів транскрипції, поява яких узгоджена з регу-
ляцією клітинного циклу: вони з'являються тільки на пізній G1-стадії,
коли й може розпочатися перемикання типів клітин.
Генетика
164
MATα
α1
αsg
asg
hsg
α2
α2 α1
MATa
αsg
asg
hsg
a1
MATα
αsg
asg
hsg
α2
α2
a1
MATa
a1
a1
α2
a1
α2
α
-клітини
а-клітини
Д
иплоїдні клітини
Рис. 5.12. Схема регуляції транскрипції в диплоїдних і гаплоїдних клітинах
двох типів S. cerevisiae. Червоні стрілки – мРНК, овали –
відповідні білкові продукти, які є факторами транскрипції груп генів,
котрі специфічно виявляються в гаплоїдних клітинах узагалі (hsg),
а-клітинах (asg) та
α-клітинах (αsg)
Але регуляція активності гена НО є ще складнішою та не до кінця
з'ясованою. Справа в тому, що після мітозу гаплоїдна спора продукує
два типи клітин, які дещо розрізняються за властивостями, – так зва-
ні материнську та дочірню. Перемикання типів клітин відбувається
тільки в материнській клітині, дочірня не здатна це робити, оскільки
ген НО не активується. Отже, материнська клітина змінює свій тип –
наприклад, а на α, і відбувається поділ з утворенням двох α-клітин,
одна з яких знову є материнською і змінює свій тип на а, а інша – до-
чірньою, вона залишається клітиною α-типу.
Така асиметрія продуктів мітозу є характерною також для дифе-
ренціації клітин багатоклітинних організмів , коли стовбурова клітина
дає початок стовбуровій клітині та клітині, котра є більш спеціалізо-
ваною та такою, що втратила частину потенціалу розвитку. Розгляну-
ті приклади відображають лише невелику частину вивчених для
S. cerevisiae генетичних механізмів, які допомагають з'ясувати загальні
закономірності функціонування еукаріотичного апарату спадковості.

Розділ 5. Генетика бактерій, вірусів і одноклітинних еукаріотів
165
Інфузорія
Генетичний апарат інфузорій відрізняється унікальною особливіс-
тю: у клітині присутні не одне, а два ядра – мікро- та макронуклеус.
Перший із них містить диплоїдний набір хромосом (наприклад,
у Tetrahymena thermophila п'ять пар хромосом), які є тільки сховищем
спадкової інформації – гени мікронуклеуса не експресуються. У мак-
ронуклеусі хромосомний набір є багатократно дуплікованим (кілька
сотень хромосом), і саме гени макронуклеуса активно експресуються,
але при цьому не передаються нащадкам.
При нестатевому розмноженні поділ мікронуклеуса здійснюється
шляхом мітозу, а макронуклеуса – шляхом простого поділу, тобто з ча-
сом він старіє, і його активність знижується. Тоді між двома клітина-
ми проходить статевий процес: відбувається кон'югація, під час якої
макронуклеуси руйнуються, мікронуклеуси розділяються шляхом ме-
йозу, і клітини обмінюються гаплоїдними ядрами – деталі процесу
можуть розрізнятися для різних видів, але в результаті утворюються
клітини, що мають по одному диплоїдному мікронуклеусу. Негайно
після цього здійснюється його мітотичний поділ, і одне з дочірніх ядер
перетворюється на макронуклеус.
Під час формування макронуклеуса геном не тільки багатократно
дуплікується, а й піддається суттєвим перебудовам. Спочатку в складі
ядра-попередника макронуклеуса проходять багатократні раунди ре-
плікації ДНК – політенізація хромосом. Далі видаляються численні
елементи послідовності ДНК (IES – internal eliminated sequences), роз-
ташовані всередині генів мікронуклеуса (таким чином, гени мікрону-
клеуса, що перериваються IES, у принципі не можуть бути експресо-
ваними). Після стикування кодуючих послідовностей генів здійсню-
ється фрагментація хромосом на своєрідні мікрохромосоми – кожна
має один або кілька генів і теломери на кінцях, які синтезуються те-
ломеразою. Під час фрагментації хромосом видаляються також усі
міжгенні зони, послідовності, що повторюються, і мобільні елементи –
прибирається все беззмістовне "сміття". Загалом видаляється до 90 %
геному. Нарешті, мікрохромосоми ампліфікуються в 4–6 раундах ре-
плікації – процес дозрівання макронуклеуса завершується.
Геном макронуклеуса інфузорії Tetrahymena thermophila вже встано-
влено. Він містить 27 тис. генів – стільки ж, скільки у вищих еукаріотів
(навіть трохи більше, ніж у людини). Унаслідок видалення значної час-
тини ДНК загальний розмір геному (гаплоїдного набору) дорівнює
105 млн пар основ (у 30 разів менше, ніж у ссавців), вміст послідовнос-
тей, що повторюються становить ~2 %. Цікавою особливістю генетично-

Генетика
166
го апарату Tetrahymena є те, що кодони UAA та UAG (стоп-кодони для
більшості організмів) кодують амінокислоту глутамін: тільки UGA вико-
ристовується як стоп-кодон.
Незвичайна система функціонування спадкового апарату інфузо-
рій ставить важливе фундаментальне питання: чому в еукаріотичних
геномах зберігається велика кількість беззмістовної ДНК? При тому,
що загальна структура геному інфузорії практично не відрізняється
від такої вищих еукаріотів, реалізується механізм видалення беззміс-
товних послідовностей при утворенні макронуклеуса. Отже, ці послі-
довності не потрібні для експресії генетичної інформації. Проте ці
"зайві" послідовності, як і в інших еукаріотів, ретельно зберігаються
в мікронуклеусі й передаються нащадкам. У чому може полягати ада-
птивне значення такого збереження, залишається не зовсім зрозумі-
лим. Утім, принаймні одна відповідь на це питання здається очевид-
ною: беззмістовна ДНК є саме тією "обкладинкою", в якій зберігається
змістовна ДНК, або своєрідним буфером, що захищає змістовний ге-
нетичний матеріал від пошкоджуючих впливів. Адже зовнішні мута-
генні фактори імовірніше спрямовані на беззмістовну ДНК через те,
що вона становить переважну більшість еукаріотичного геному.
Контрольні запитання і завдання
1. Яка різниця між бактеріальними штамами типів F
+
і Hfr?
2. Якими шляхами здійснюється перенесення генетичного матеріалу
між бактеріями?
3. На які класи можна поділити бактеріофаги за типом носія генетич-
ної інформації?
4. Поясніть різницю між сайт-специфічною та гомологічною рекомбінаці-
єю. Яке функціональне значення має сайт-специфічна рекомбінація?
5. Опишіть життєвий цикл бактеріофага λ. Від чого залежить вибір
шляхів розвитку цього фага?
6. Яку функцію виконує репресор фага λ? У чому полягає механізм йо-
го дії?
7. На які класи та за яким принципом поділяють еукаріотичні віруси?
8. Опишіть цикл розвитку ретровірусів.
9. Як відбувається розмноження Saccharomyces cerevisiae?
10. За яким механізмом здійснюється перемикання типів спарювання
в S. cerevisiae?
11. У чому полягає структурна та функціональна різниця між макро-
та мікронуклеусом у інфузорій?

РОЗДІЛ 6
Генетика
багатоклітинних еукаріотів
Найхарактернішими особливостями організації спадкового апара-
ту еукаріотів порівняно з прокаріотами є (див. також розділи 1 і 2):
• Наявність клітинного ядра з кількома молекулами ДНК, які є ком-
понентами складних білково-нуклеїнових комплексів – хромосом.
• Присутність, поряд із головним ядерним спадковим апаратом,
цитоплазматичних елементів спадковості – ДНК усередині мі-
тохондрій і хлоропластів.
• Великий розмір еукаріотичних геномів, причому значна частка
припадає на послідовності ДНК, які повторюються багато разів.
• Мозаїчний принцип будови еукаріотичних генів, причому один
ген може давати кілька різних функціональних продуктів за
рахунок альтернативного сплайсингу.
• Велика кількість генів, причому всі клітини багатоклітинного
організму (за винятком деяких) містять ідентичні набори генів,
хоча більша їхня частка є неактивною у клітинах даного типу.
Це викликає необхідність реалізації складної системи регуляції
експресії генів у процесі диференціювання клітин під час онто-
генезу. Одним із важливих елементів цієї системи є епігенетич-
на спадковість – забезпечення спадкування дочірніми клітина-
ми не просто набору генів, а й системи специфічного блоку-
вання певної їхньої частини.
• Статевий процес, що лежить в основі розмноження переважної
більшості багатоклітинних організмів: нащадок отримує два
набори алельних генів від двох батьків різної статі. Механізми
визначення статі є важливим розділом генетики еукаріотів.
Базуючись на матеріалі попередніх розділів, розглянемо ці особли-
вості детальніше.
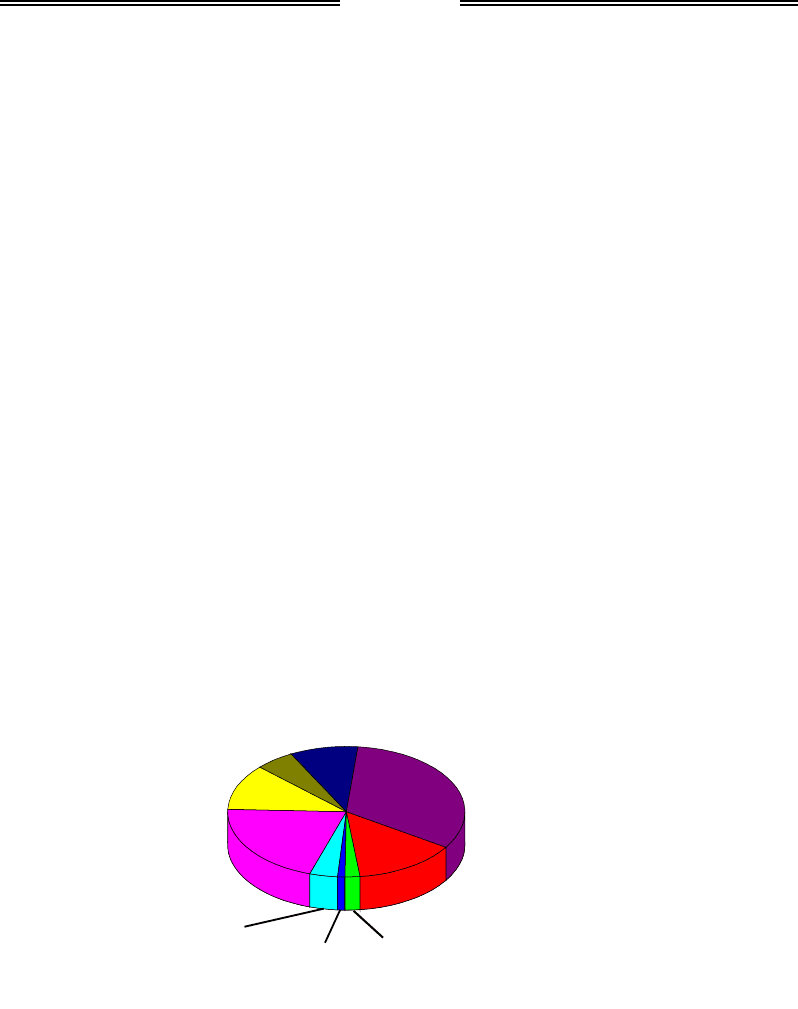
Генетика
168
ЕУКАРІОТИЧНІ ГЕНОМИ
Загальні риси будови еукаріотичних геномів
Усі гени багатоклітинного організму можна розділити на дві групи:
1) гени, від яких залежать певні універсальні функції, і які є актив-
ними в усіх клітинах, – "гени домашнього господарства" (housekeeping
genes); 2) гени, що специфічно активуються в клітинах певного типу, –
"гени розкоші " (luxury genes). Досить загальною ознакою генів пер-
шої групи є розташування в їхніх регуляторних зонах так званих
CpG-острівців – ділянок із підвищеним вмістом динуклеотидів CpG
(мається на увазі послідовність нуклеотидів уздовж подвійної спіралі).
Загалом вміст цих динуклетидів у еукаріотичних геномах приблизно
у п'ять разів менший за очікуваний унаслідок метилювання цитозину
в складі CpG-контакту: 5mC (5-метилцитозин) спонтанно перетворю-
ється на тимін, що є одним із джерел виникнення мутацій. Метил-
ування цитозину в регуляторних ділянках є одним із механізмів реп-
ресії генів (див. нижче). Відповідно, гени, які зберігають свою актив-
ність у більшості клітин, містять неметильовані динуклеотиди CpG,
вміст яких зберігається на високому рівні.
Загальна кількість білкових генів у геномах вищих еукаріотів ва-
ріює приблизно від 20 до 30 тис. (див. табл. 1.1). Приблизний розподіл
еукаріотичних білків за їхніми функціями показано на рис. 6.1.
Цитоскеле
т
Транспорт
Інші + невідомі
Взаємодія з
нуклеїновими кислотами
Регуляція клітинного циклу
Фактори з
б
ірки
білкових структур
Захист / імунна
система
Ферменти
Сигнальна
т
рансдукція
Рис. 6.1. Приблизний розподіл
білків еукаріотичного протеому за функціями

Розділ 6. Генетика багатоклітинних еукаріотів
169
Серед еукаріотичних генів 25–50 % є унікальними (представлені
в геномі єдиною копією), решта належать до родин генів, що склада-
ються з кількох копій, як правило, не ідентичних. Відповідні (гомоло-
гічні але не ідентичні) білки становлять родину білків. Кілька родин
(протеїнкінази, транскрипційні фактори певного типу, імуноглобулі-
ни) містять сотні білків, більшість родин складається з кількох (до 30)
білків. Гени такої родини часто об'єднані в геномі в кластери – знахо-
дяться поряд у певній хромосомі (кластери генів теплового шоку, гло-
бінові гени). Слід зауважити, що такий кластер не є опероном – кожен
ген піддається регуляції як окрема одиниця транскрипції. Наприклад,
кластер генів β-субодиниці гемоглобіну містить гомологічні гени, які
активуються на певних стадіях індивідуального розвитку (рис. 6.2).
Проте всі гени кластеру мають також спільну регуляторну зону, яка
відповідає за загальний потенційно активний стан кластеру в кліти-
нах, які в принципі мають здійснювати синтез гемоглобіну.
εγγψβδβ
~60 000 пар основ
Ембріон
Утро
б
ний
розвиток
Псевдоген
Д
орослий
організм
Рис. 6.2. Кластер β-глобіну в 16 хромосомі людини
(кожен ген містить інтрони). Указано стадії розвитку,
на яких відповідні гени є активними
β-Глобіновий кластер містить також неактивний псевдоген (див.
розділ 1). Після дезактивації гена (порушення внаслідок мутації іні-
ціації транскрипції, сплайсингу тощо) псевдоген перестає бути об'-
єктом відбору, і в ньому накопичується велика кількість мутацій.
Зрозуміло, що в першу чергу псевдогени з'являються саме в класте-
рах – коли є кілька копій гена , і пошкодження од ного з них не при-
водить до фатальних наслідків.
Кілька генних кластерів повторюються в геномі багато разів. Серед
білкових генів це стосується генів гістонів (див. розділ 1) – гени п'яти мо-
лекул гістонів завжди згруповані в кластер (кожен ген – окрема одиниця
транскрипції), який повторюється до 100 разів. Іншим прикладом клас-
терів, що повторюються, є гени рибосомної РНК (див. розділ 2), але
в цьому випадку кластер є одиницею транскрипції.
Крім генів, що повторюються, еукаріотичний геном містить значну
кількість інших послідовностей, що повторюються (до 50 % геному).
Основні типи таких повторів, крім уже згаданих псевдогенів, такі:

Генетика
170
1. Тандемні повтори. До таких відносять багатократні повтори
коротких послідовностей по 6–8 пар основ у теломерах і повтори так
званої α-сателітної ДНК у центромерах (довжина повтору варіює від
7 пар основ у дрозофіли до 100–200 пар основ у ссавців, у людини –
171 пара основ). По всьому геному розподілені також так звані прості
повтори (SSR, simple sequence repeats). Зазвичай виділяють мікроса-
теліти – 1–15 пар основ, що повторюються від 10 до кількох тисяч
разів, і мінісателіти – 15–500 пар основ, що повторюються до
100 разів. Кількість міні- та мікросателітних локусів становить десят-
ки й сотні тисяч. Розподіл локусів за кількістю повторів є специфіч-
ною індивідуальною ознакою – на кшталт відбитків пальців.
2
. Сегментні дуплікації – великі блоки довжиною 1–200 тис. пар
основ, які характеризуються високим ступенем гомології. Імовірно,
сегментні дуплікації є продуктом колишніх порушень хромосом. Час-
тіше зустрічаються в перицентромерних і субтеломерних зонах.
3.
Інтерсперсні (мобільні) елементи, здатні до переміщення та
розмноження в межах геному, становлять основну кількість ДНК, що
повторюється. Частина таких послідовностей є результатом колиш-
ньої активності мобільних елементів (таких, що втратили здатність
до переміщення). Основні типи еукаріотичних мобільних елементів:
•
ДНК-транспозони – переміщення здійснюється шляхом вирі-
зання ділянки ДНК із наступним вбудовуванням її в інше місце –
цілком аналогічно до відповідних елементів у прокаріотів.
Транспозони містять один або два гени (у різних видів), що ко-
дують транспозазу – фермент, який забезпечує транспозицію
елемента, – його вирізання з донорного сайта та вбудовування
в сайт-мішень. Гени транспозази можуть бути пошкодженими –
тоді транспозиція даного елемента відбувається з використан-
ням транспозази, закодованої іншим ДНК-транспозоном.
Кодуюча частина транспозона фланкована невеликими ін-
вертованими повторами, які впізнає транспозаза, вирізаючи
фрагмент ДНК. Сайт-мішень – невелика специфічна послідов-
ність ДНК, котра теж упізнається транспозазою і теж розріза-
ється, після чого транспозаза каталізує вбудовування транспо-
зона до сайта-мішені (рис. 6.3). Процес транспозиції залишає
дволанцюговий розріз у місці, де містився транспозон. У разі
незалежної від реплікації транспозиції (нереплікативна транс-
позиція), цей розріз піддається репарації за механізмом негомо-
логічного з'єднання кінців (див. розділ 1). Тобто транспозон
просто "стрибає" в інше місце. Але досить часто транспозиція