Реферат - Классификация сложных белков. Гликопротеины. Лектины растительных организмов. Защитные белки
Подождите немного. Документ загружается.

МИНИСТЕРСТВО ОБЩЕГО И ПРОФЕССИОНАЛЬНОГО
ОБРАЗОВАНИЯ РФ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
ГОУ ВПО Башкирский Государственный Университет
Биологический факультет
Кафедра биохимии и биотехнологии
Реферат на тему :
«Классификация сложных белков.
Гликопротеины. Лектины растительных
организмов.
Защитные белки»
Выполнила:
студент 2-го курса
биологического факультета
группы Б: Миниярова А.Д.
Проверили: д.б.н., проф.
Ямалеева А.А.,
к.б.н., доцент Баширова Р.М.
Уфа-2007
2
Оглавление
Введение...................................................................................................................................2
Глава 1. КЛАССИФИКАЦИЯ СЛОЖНЫХ БЕЛКОВ
..........................................................................3
1.1 Липопротеины...............................................................................................................3
1.2. Фосфопротеины……………………………………………………………………….5
1.3. Металлопротеины……………………………………………………………………….5
1.4. Хромопротеины...........................................................................................................6
Глава 2. Гликрпротеины…………………………………………………………………………..8
2.1. Классификация гликопротеинов.................................................................................10
2.2. Защитные белки в плазме крови ...................................................................................11
2.2.1.Строение иммуноглобулинов………………………………………………………....13
2.2.2. Описание показателей иммунограммы .
Концентрация иммуноглобулинов классов G,A,М…………………………………18
2.3. Биологическая роль гликопротеинов............................................................................20
2.4. Лектины………………………………………………………………………………..22
2.4.1. Несколько слов об открытии лектина……………………………………………...25
2.4.2. Зачем клетке лектины………………………………………………………….........26
2.4.3. Лектины в биотехнологии…………………………………………………………...28
ЗАКЛЮЧЕНИЕ......................................................................................................................34
3
Введение.
В медицине успехи биохимии определяют стратегию создания и применения
лекарственных веществ, являются источником новых методов для диагностики заболеваний и
основой для поразительных открытий, касающихся выяснения причин тех или иных
патологических процессов в организме. В частности, оказалось, что многие болезни,
предающиеся по наследству, возникают в результате нарушения отдельных звеньев обмена
белков, углеводов, липидов, нуклеиновых кислот, гормонов. Это послужило причиной для
выделения в отдельную ветвь медицины учения о молекулярных основах патологии и, в
частности, об энзимопатиях, т. е. Нарушениях функций ферментов, приводящих к развитию
заболеваний.
Применение многочисленных и разнообразных химических препаратов в
животноводстве и растениеводстве, базирующееся на данных биохимии и физиологии,
способствует подъему продуктивности указанных отраслей сельского хозяйства и
производительности труда.
Всестороннее изучение биохимии микроорганизмов и открывшиеся при этом широкие
перспективы их практического использования привели в нашей стране к созданию
промышленности микробиологического синтеза вместо разрозненных бродильных
производств. Ее продукцию составляет кормовой белок (пищевой белок), аминокислоты, все
без исключения антибиотики (кормовые и медицинские), многие витамины и практически все
гормоны и ферменты.
Русские и советские ученые внесли большой вклад в развитие биохимии.
Основоположником отечественной биохимии по праву считают А.Я. Данилевского (1839-
1923). Он впервые высказал идею об обратимости действия биологических катализаторов –
ферментов и, исходя из этого, осуществил синтез белковоподобных веществ – пластеинов.
Занимаясь изучением белков, А.Я. Данилевский предложил, что составляющие их
структурные единицы соединены друг с другом пептидными связями.
4

Глава 1. МНОГООБРАЗИЕ БЕЛКОВ
Белки играют решающую роль в жизнедеятельности отдельных клеток и всего
многоклеточного организма, так как их функции удивительно многообразны. Многообразие функций
белков определяется особенностями их первичной структуры и конформации, уникальностью
строения активного центра и способностью связывать специфические лиганды. Лишь очень
небольшая часть всех возможных вариантов пептидных цепей может принять стабильную
пространственную структуру; большинство из них может принимать множество конформации с
примерно одинаковой энергией Гиббса, но с различными свойствами. Первичная структура известных
белков, отобранных биологической эволюцией, обеспечивает ис
ключительную стабильность одной из
конформации,
которая определяет особенности функционирования этого белка.
1.1 Гомологичные белки
Гомологичными называют белки, выполняющие у разных видов одинаковые функции, например гемо-
глобин у всех позвоночных осуществляет транспорт 0
2
, цитохром с - митохондриальный белок, участ-
вующий в процессах биологического окисления.
Гомологичные белки большинства видов:
- имеют одинаковую или очень близкую молекулярную массу;
- во многих положениях содержат одни и те же аминокислоты, называемые инвариантными остатками;
- в некоторых положениях наблюдаются значительные различия аминокислот - так называемые
вариабельные аминокислотные остатки;
- содержат гомологичные последовательности — совокупность сходных черт в аминокислотной по-
следовательности сравниваемых белков (кроме идентичных аминокислот, эти последовательности
содержат разные, но близкие по физико-химическим свойствам аминокислотные радикалы). Цитохром
с — митохондриальный белок, участвующий в биологическом окислении. Установлена его
5

аминокислотная последовательность более чем у 60 видов. В 27 положениях находятся инвариантные
аминокислотные остатки. Цитохромы с дрож
жей и лошади различаются по 48 аминокислотным
остаткам, курицы и утки - по 2 аминокислотам, курицы и индейки идентичны.
Сравнение аминокислотной последовательности гомологичных белков выявило:
1) консервативные, инвариантные аминокислотные остатки важны для формирования уникальной
пространственной структуры и биологической функции данных белков;
2) наличие гомологичных белков говорит об общем эволюционном происхождении видов;
3) число вариабельных аминокислотных остатков в гомологичных белках пропорционально
филогенетическим различиям между сравниваемыми видами;
4) в некоторых случаях даже небольшие изменения аминокислотной последовательности могут
вызвать нарушения свойств и функций белков;
5) но далеко не все изменения аминокислотной последовательности вызывают нарушения биоло-
гических функций белков;
6) наибольшие нарушения структуры и функции белков возникают при замене аминокислот:
— входящих в ядро сворачивания (набор аминокислот, с которых начинается формирование кон-
формации);
— входящих в состав активного центра;
— на участках пересечения полипептидной цепи при образовании третичной структуры.
Серповидно-клеточная анемия — заболевание, при котором в крови больного вместо НbА обна-
руживается НbS. В его β-цепи в положении 6 вместо Глу стоит Вал. Наличие на поверхности
НbS валина приводит к агрегации молекул дезокси-НbS, образованию нерастворимых нитей
и деформации эритроцитов, которые часто приобретают форму серпа.
Почти все замены аминокислот, обнаруженные на поверхности НЬA, безвредны; НЬS —
поразительное исключение.
1.2. Семейства белков
В ходе эволюции в пределах одного биологического вида замены аминокислотных остатков могут
приводить к возникновению разных белков, выполняющих родственные функции и имеющих
гомологичные последовательности аминокислот.
Они имеют поразительно сходные конформации: количество и взаиморасположение ос-спиралей
и/или β-структур, большинство поворотов и изгибов полипептидных цепей похожи или
идентичны.
6
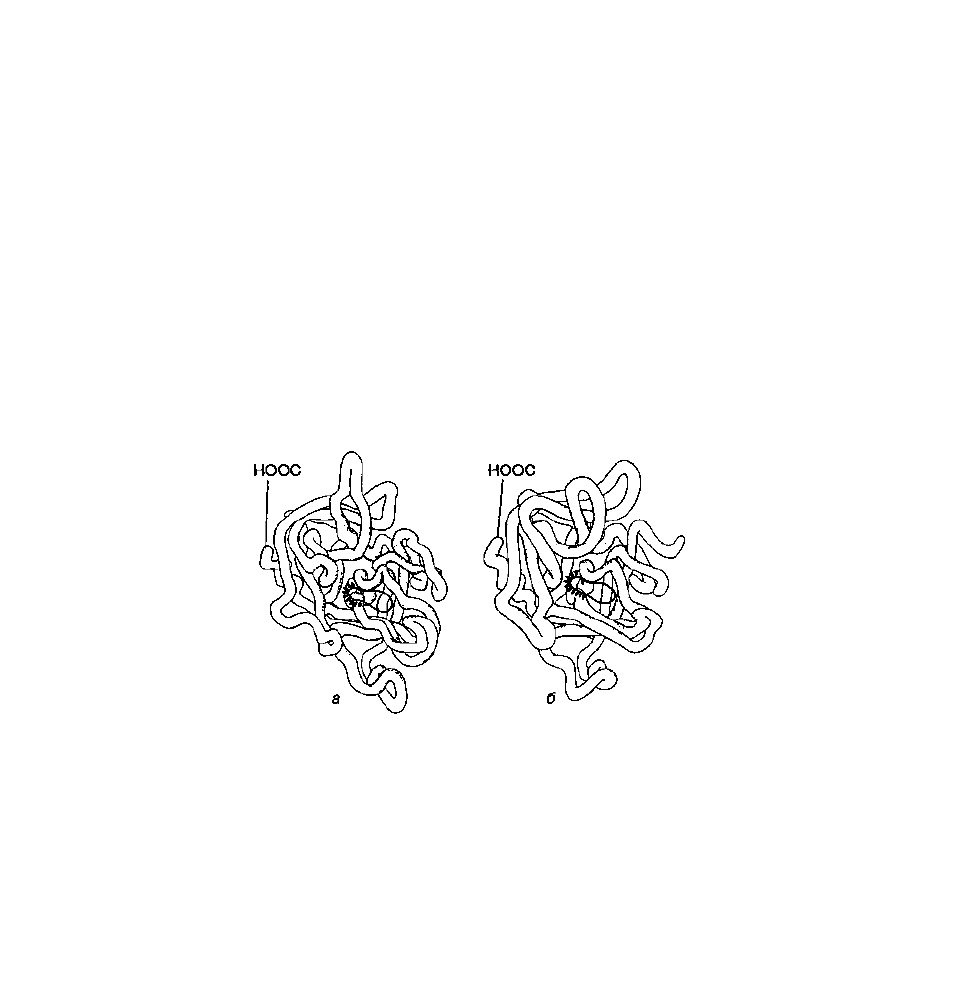
Белки, имеющие гомологичные участки полипептидной цепи, сходную конформацию и родст-
венные функции, выделяют в семейства белков. Примеры семейств белков:
- сериновые протеиназы;
- семейство иммуноглобулинов;
- семейство миоглобина.
Сериновые протеиназы - семейство белков, выполняющих функцию протеолитических ферментов.
К ним относятся пищеварительные ферменты - химотрипсин, трипсин, эластаза и многие
факторы свертывания крови . Эти белки имеют в 40% положений идентичные аминокислоты и очень
близкую конформацию (рис.1).
Рис. 1. Пространственные структуры эластазы (а) и химотрипсина (б).
Некоторые аминокислотные замены привели к изменению субстратной специфичности
этих белков и возникновению функционального многообразия внутри семейства.
7

Глава 2. КЛАССИФИКАЦИЯ БЕЛКОВ
1. По форме молекул белки можно разделить на две большие группы - глобулярные (имеющие
сферическую форму) и фибриллярные (удлиненной формы).
2. По наличию или отсутствию в белке неаминокис
лотной части
они делятся на
простые
(
состоящие
только из аминокислот) и
сложные
(
имеющие в своем составе компонент неаминокислотной природы).
3. По функциям, выполняемым белками, их можно разделить на структурные, сократительные
транспортные, каталитические, защитные, рецепторные, регуляторные и др.
Полипептидную часть сложного белка называют апопротеином, неаминокислотный компонент -
простетической группой, а весь белок - холопротеином.
В каждую из этих функциональных групп входит огромное количество индивидуальных белков.
2.1. Суперсемейство иммуноглобулинов
В работе иммунной системы огромную роль играют белки суперсемейства иммуноглобулинов, кото-
рое включает в себя 3 семейства белков: антитела (иммуноглобулины), рецепторы Т-лимфоцитов,
белки главного комплекса гистосовместимости - МНС 1-го и 11-го классов (major histocompatibility
complex).
Все они имеют доменное строение, состоят из гомологичных иммуноподобных доменов и вы-
полняют сходные функции — взаимодействуют с чужеродными структурами, либо
растворенными в крови, лимфе или межклеточной жидкости (антитела), либо находящимися
на поверхности клеток (собственных или чужеродных).
Антитела — специфические белки, вырабатываемые В-лимфоцитами в ответ на попадание в орга-
низм чужеродной структуры, называемой антигеном.
Особенности строения антител
1. Простейшие молекулы антител состоят из 4 полипептидных цепей: 2 идентичных легких
— L,содержащих около 220 аминокислот и 2 идентичных тяжелых — Н, состоящих из 440-
700 аминокислот. Все 4 цепи антитела соединены множеством нековалентных и 4
дисульфидными связями.
2. Легкие цепи антитела состоят из 2 доменов: вариабельного, находящегося на N-концевой
области полипептидной цепи и константного, расположенного на С-конце.
3. Тяжелые цепи обычно имеют 4 домена: один вариабельный, находящийся на N-конце и 3
константных.
8

4. Каждый домен иммуноглобулина имеет β-складчатую суперструктуру, в которой 2
остатка цистеина соединены дисульфидной связью.
5. Между 2 константными доменами
имеется участок, содержащий большое количество остатков
пролина, которые препятствуют форми
рованию вторичной структуры и взаимодействию соседних
Н-цепей на этом отрезке. Эта шарнирная область придает молекуле антитела гибкость.
6. Между вариабельными доменами тяжелых и легких цепей находятся 2 идентичных
антигенсвязывающих участка, поэтому такие антитела часто
называют бивалентами.
В связывании антигена с антителом участвует не вся аминокислотная последовательность
вариабельных участков обеих цепей, а всего лишь 20—30 аминокислот, расположенных в
гипервариабельных областях каждой цепи. Именно эти области определяют уникальную
способность каждого вида антетата взаимодействовать с соответствующим комплементарным
антигеном.
Антитела - одна из линий защиты организма против внедрившихся чужеродных организмов. Их
функционирование можно разделить на два этапа:
первый этап - узнавание и связывание антигена на
поверхности чужеродных организмов, которые происходят благодаря наличию в структуре антите-
ла антигенсвязывающих участков; второй этап — инициация процесса, благодаря которому антиген
инактивируется и разрушается. Специфичность вто
рого этапа зависит от класса антител.
Существует 5 классов тяжелых цепей, отличаю
щихся по строению константных доменов: α, δ, ε, γ и μ, в
соответствии с которыми различают 5 классов иммуноглобулинов: A, D, E, G и М.
Особенности строения тяжелых цепей придают шарнирным участкам и С-концевым областям тя-
желых цепей характерную для каждого класса конформацию.
После связывания антигена с антителом конформационные изменения константных доменов
определяют путь удаления антигена в организме.
Иммуноглобулины М.
Иммуноглобулины М имеют две формы:
Мономерная форма - первый класс антител, продуцируемый развивающимся В-
лимфоцитом. Впоследствии многие В-клетки переключаются на выработку других классов
антител, но с тем же антигенсвязывающим участком.
1gМ встраивается в мембрану и выполняет роль антигенраспознающего рецептора.
Встраивание 1gМ в мембрану клеток возможно благодаря наличию в хвостовой части
участка 25 гидрофобных аминокислотных остатков.
Секреторная форма 1gМ содержит 5 мономерных субъединиц, связанных друг с другом
дисульфидными связями и дополнительной полипептидной J-цепью (рис. 2.1).
9
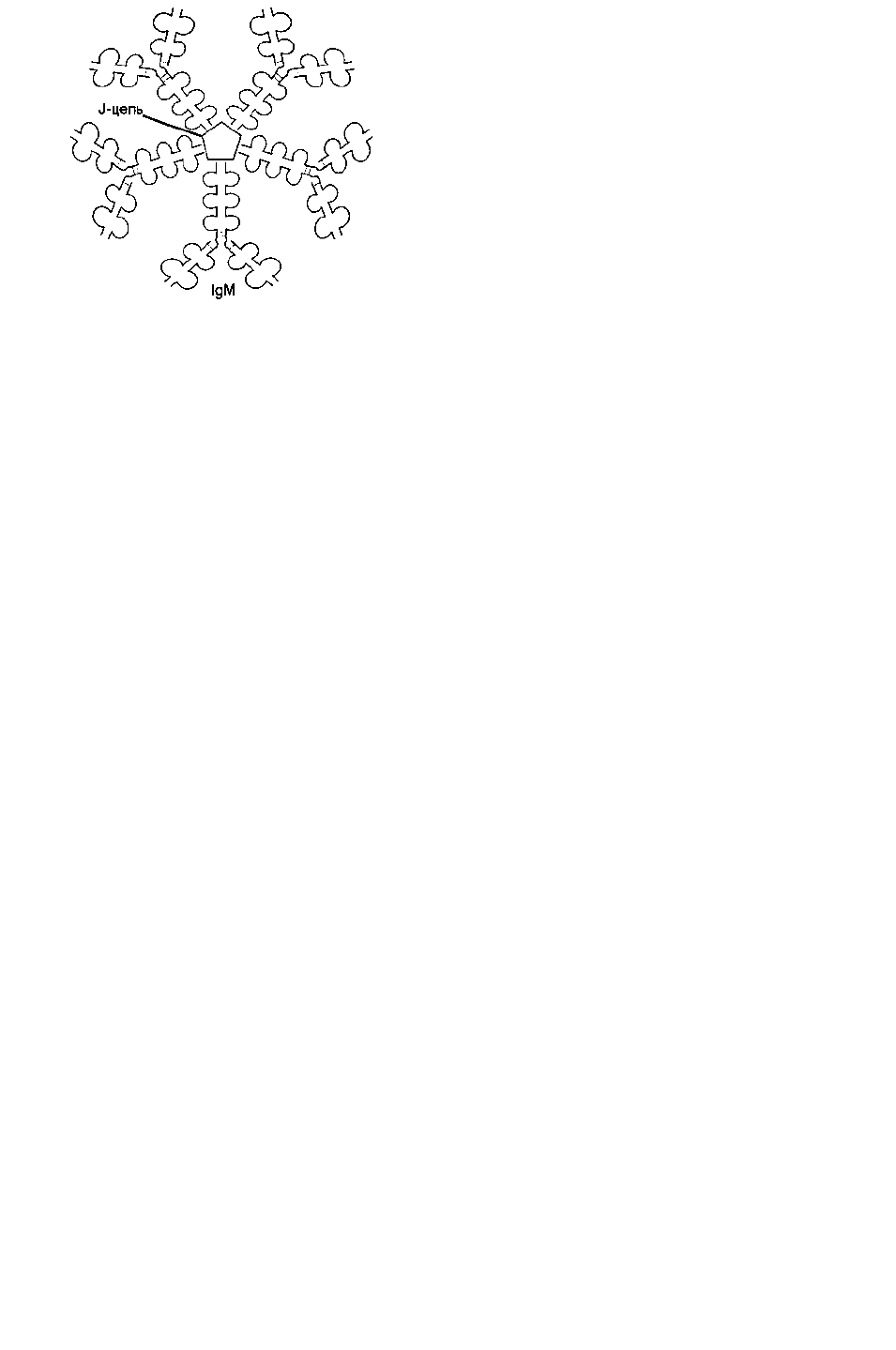
Рис. 2.1. Строение секреторной формы 1gМ.
Тяжелые цепи мономеров этой формы не содержат гидрофобной хвостовой части. Пентамер имеет
10 центров связывания с антигеном.
Секреторная форма 1gМ — основной класс антител, секретируемых в кровь при первичном им-
мунном ответе.
Связывание 1gМ с антигеном изменяет конформацию 1gМ и индуцирует связывание его с первым
компонентом системы комплемента и активацию этой системы. Если антиген расположен на
поверхности микроорганизма, система комплемента вызывает нарушение целостности клеточной
мембраны и гибель бактериальной клетки.
Иммуноглобулины G.
В количественном отношении этот класс иммуноглобулинов доминирует в крови (75% ).
1gG — мономеры, основной класс антител, секретируемый в кровь при вторичном
иммунном ответе. После взаимодействия 1gG с поверхностными антигенами
микроорганизмов комплекс антиген—антитело:
— способен связывать и активировать белки системы комплемента;
— может взаимодействовать со специфическими рецепторами макрофагов и нейтрофилов,
что приводит к фагоцитозу комплексов антиген—антитело и разрушению их в фагосомах;
— 1gG — единственный класс 1g, способный проникать через плацентарный барьер и
обеспечивать внутриутробную защиту плода от инфекций.
Иммуноглобулины D
Иммуноглобулины D обнаружены в сыворотке в очень небольшом количестве, являются
мономерами. В тяжелых δ-цепях имеют 1 вариабельный и 3 константных домена. Выполняют
роль рецепторов В-лимфоцитов; другие функции пока неизвестны.
Взаимодействие специфических антигенов с рецепторами на поверхности В-лимфоцитов приво-
дит к передаче этих сигналов в клетку и включению механизмов, обеспечивающих размножение
данного клона лимфоцитов.
10