Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.


are bound to the telomere via
the same Dro-
tein, Cdcl3:
.
The Stnl
protein protects
against
de-
gradation (specifically,
against
,any
ex-
tension
of the degradation
of the
C-A-strand
that
generates
the G-tail).
.
A telomerase
enzyme extends the C-
A-rich strand. Its
activity is influenced
by two
proteins
that have
ancillary
roles,
such as controlling
the
lengttL
of the
extension.
The
telomerase uses the 3'-OH
of the G+T
telomeric strand
as a
primer
for synthesir; of tan-
dem TTGGGG repeats.
Only dGTP and dTTP
are
needed for the activity. The
telomerase irs a large
ribonucleoprotein
that consists of a templating
RNA
(coded
by TLCl) and
a
protein
with cat-
alytic activity
(EST2)
The short RNA
,rompo-
nent
(159
bases long in Tetrahymena,
and 192
bases long in Euplotes) includes
a sequence of 1 5
to 22 bases that is identical
to two repeats of the
C-rich repeating
sequence. This RNA
F,rovides
the template for synthesizing
the G-rich
repeat-
ing
sequence.
The
protein
component of the
telomerase
is
a catalytic subunit
that can act
only upon the RNA template
provideclby
the
nucleic acid component.
i'ri. r :
i::i:' .:.i
;i i
shows the action of telomerase.
The enzyme
progresses
discontinuou:;ly: The
template RNA
is
positioned
on the DNA
primer,
several nucleotides are added
to the
prin:rer,
and
then the enzyme translocates
to begin again.
The
telomerase
is
a specialized example of a
reverse transcriptase, an enzyme that
synthe-
sizes
a DNA sequence
using an RNA template
(see
Section 22.4, Yiral DNA Is
Generated
by
Reverse Transcription). We do not know how
the complementary
(C-A-rich)
stran<i of the
telomere is assembled,
but we may spreculate
that it could be synthesized by using th,.' 3'-OH
of a terminal G-T
hairpin
as a
primer
for DNA
synthesis.
Telomerase
synthesizes the inclividual
repeats
that are added to the chromosome ends,
but does
not itself
control the number of repeats.
Other
proteins
are involved in determining the
length
of
the telomere. They
can be
identified
by the
ESTI
and
ESTJ mutants
in
yeast
that have
altered telomere
lengths.
These
proteJLns
may
bind telomerase, and influence the le ngth of
the telomere by controlling the access of telo-
merase to its substrate. Proteins that bind telo-
meres in mammalian cells have
been
found,
but less is known about their functionrs.
Each organism has a characterisic range of
telomere lengths. They are long in mammals
i:Li:li-:iii.
iii .i i Tetomerase
positions
itsetf by base
pair-
ing between the
RNA template
and the
protruding
single-
stranded
DNA
primer.
It
adds
G and
T bases one
ata time
to the
primer,
as directed
by the
template.
The cycle starts
again when one
repeating unjt
has been
added.
(typically
5 to l5 kb
in human
beings) and
short
in
yeast
(typically
-300
bp in S.
cerevisiae).
The
basic control
mechanism
is
that the
probability
that a telomere will
be
a substrate
for telo-
merase
increases as
the
length of the
telomere
shortens; we
do not
know
if this
is
a
continu-
ous effect or
if it depends
on the
length
falling
below some critical
value.
When
telomerase
acts on
a telomere,
it may
add several
repeat-
ing units.
The enzyme's
intrinsic
mode
of action
is to dissociate
after adding
one
repeat;
addition
of several repeating
units
depends
on other
pro-
teins
that cause telomerase
to undertake
more
than one
round of extension.
The
number of
repeats that
is
added
is not
influenced
by the
length of the
telomere
itself,
but instead
is con-
trolled by ancillary
proteins that associate
with
telomerase.
Binding: RNA template
pairs
with DNA
primer
DNA
primer
3'W
5:
3',
RNA templal
Polymerization:
RNA template direc
of nucleotides to 3'end
of DNA
Polymerizationcontinues
I
to end of template
region
Y
J
3',
Translocalio;r:
Enzyme moves
to
5',..TTGGGGTTGGGGTTGC
3',
.,.AACCCC 5',
MCCC
3',
28.18
Tetomeres
Are Synthesized
by a Ribonucleoprotein
Enzyme 757
The
minimum features required
for exis-
tence as a
chromosome are:
.
Telomeres
to ensure
survival.
o
A centromere to
support segregation.
.
An origin to initiate
replication.
All of these
elements have been
put
together
to construct
a
yeast
artificial chromosome
(YAC).
This is
a useful method for
perpetuating
foreign
sequences.
It turns
out that the synthetic
chro-
mosome
is stable
only
if
it is longer
than
20
to
50 kb.
We do not know
the basis for
this effect,
but the ability
to construct a
synthetic chromo-
some
allows us to investigate
the nature
of the
segregation
device in a controlled
environment.
@
Telomeres
Are Essential
for
SurvivaI
Telomerase
activity
is found in
all dividing cells
and is
generally
turned
off
in
terminally differ-
entiated
cells that do not
divide.
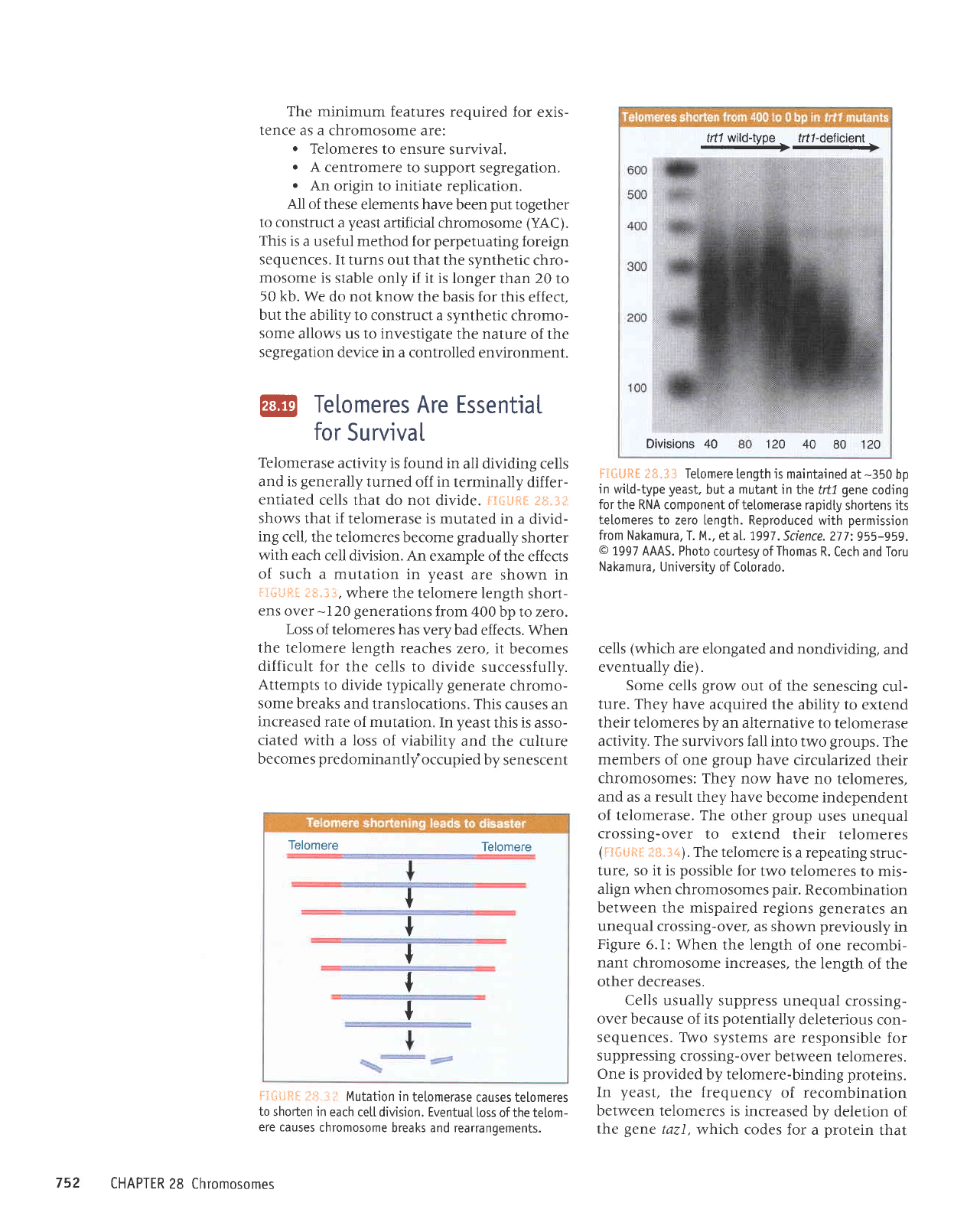
F{{;i.ffts
I*.3f
shows
that if telomerase is
mutated in a
divid-
ing
cell, the telomeres
become
gradually
shorter
with each
cell division. An
example
of the effects
of such a mutation
in
yeast
are shown in
lii;Lsgtt
IS.i3,
where the
telomere length
short-
ens
over
-
I 20
generations
from 400
bp to zero.
Loss
of telomeres
has very
bad effects. When
the
telomere length reaches
zero, it
becomes
difficult
for
the cells to
divide successfully.
Attempts
to divide typically
generate
chromo-
some
breaks and translocations.
This
causes an
increased
rate of mutation.
In
yeast
this is asso-
ciated
with a loss
of viability
and the culture
becomes
predominantly'occupied
by senescent
i:i$11*il
;.18.-iii
Mutation in
tetomerase
causes telomeres
to
shoden in
each cetl division. Eventual
loss
ofthe telom-
ere causes
chromosome
breaks and rearranqements.
CHAPTER
28 Chromosomes
f,ttwild-tvpe frtT-deficient
Divisions
40 120 40
80 120
ijil'tjEtil f
S.S.3
Tetomere
length
js
maintained
at
-350
bp
in
wiLd-type
yeast,
but a
mutant
in the
frfl
gene
coding
for
the RNA component oftelomerase
rapidty
shortens its
telomeres
to zero Length. Reproduced
with
permission
from Nakamura,
T. M., et a|.1,997.
Science. 277:955-959.
o
1997
AAAS. Photo
courtesy of
Thomas
R.
Cech and Toru
Nakamura, University
of Co[orado.
cells
(which
are elongated
and nondividing,
and
eventually die).
Some
cells
grow
out of the
senescing
cul-
ture. They
have acquired
the ability to
extend
their
telomeres by an alternative
to telomerase
activity. The survivors
fall into two
groups.
The
members
of one
group
have
circularized
their
chromosomes:
They now have
no telomeres,
and
as a
result
they have
become independent
of telomerase. The
other
group
uses
unequal
crossing-over
to extend
their telomeres
(ft**ftf
f *-34). The telomere
is a repeating
struc-
ture, so it is
possible
for two telomeres
to mis-
align when chromosomes
pair.
Recombination
between the mispaired
regions
generates
an
unequal crossing-over,
as shown
previously
in
Figure 6.1: When
the length
of one recombi-
nant chromosome
increases.
the length
of the
other decreases.
Cells usually
suppress unequal
crossing-
over because
of
its
potentially
deleterious
con-
sequences. Two
systems
are responsible
for
suppressing
crossing-over
between telomeres.
One is
provided
by telomere-binding proteins.
In
yeast,
the frequency
of recombination
between telomeres is
increased
by deletion
of
the
gene
tazl,
w};'ich codes for
a
protein
that
Telomere
Telomere
+
+
I
+
+
752

regulates
telomerase activity. The
second is a
general
system that
undertakes mismatch repair.
In
addition to correcting mismatched
base
pairs
that may arise in DNA,
this system suppresses
recombination
between mispaired regions. As
shown in Figure 28.34, this includes
telomeres.
When
it is mutated,
a
greater
proportion
of
telomerase-deficient
yeast
survive the loss of
telomeres because recombination
between
telomeres
generates
some chromosomes with
longer telomeres.
When eukaryotic cells
are
placed
in cul-
ture, they usually divide for a fixed number
of
generations
and then
enter senescence. The rea-
son appears to be a decline in telomere length
because of the absence of telomerase
expres-
sion. Cells enter a crisis from
which some
emerge, but typically the cells that emerge have
chromosome
rearrangements
that have resulted
from
lack
of
protection
of chromosome ends.
These rearrangements may
cause
mutations
that contribute to the
tumorigenic state. The
absence of telomerase expression in this situa-
tion is due to failure to express
the
gene,
and
reactivation of telomerase is one of the mech-
anisms by
which these
cells then survive con-
tinued culture.
(This
of course was not an option
in the
yeast
experiments in which
the
gene
had
been deleted.)
Sum
mary
The
genetic
material
of all organisms and viruses
takes the form of tightly
packaged
nucleopro-
tein. Some virus
genomes
are inserted into
pre-
formed
virions, whereas
others assemble
a
protein
coat around the
nucleic
acid.
The
bac-
terial
genome
forms
a dense nucleoid, with
-20o/o
protein
by mass, but details of the inter-
action of
the
proteins
with DNA
are
not known.
The DNA is organized into
-100
domains that
maintain
independent
supercoiling, with a den-
sity of unrestrained supercoils corresponding
to
-llI00
to 200 bp. In eukaryotes, interphase
chromatin
and metaphase chromosomes both
appear to be organized
into large loops. Each
loop may be an independently supercoiled
domain.
The
bases of the
loops
are connected
to a metaphase scaffold or to the nuclear
matrix
by specific DNA sites.
Transcriptionally active sequences reside
within the euchromatin that comprises the
majority of interphase chromatin. The regions
of heterochromatin are
packaged
-5
to l0x more
compactly,
and are transcriptionally inert. AII
{- it-'
{J
ii 11 .,i
:.;.r.
-'-l
+
Crossi
n
g-ove
r i n
telo
meric
regio ns is usu-
a[[y suppressed
by mismatch-repair
systems, but
can occur
when
they are
mutated.
An unequal
crossing-over
event
extends
the
tetomere of
one of the
products.
a[[owing
the
chromosome to survive
in the absence
of
telomerase.
chromatin becomes
densely
packaged
during
cell division, when
the individual
chromosomes
can be distinguished.
The existence
of
a repro-
ducible ultrastructure
in chromosomes
is indi-
cated by the
production
of
G-bands
by treatment
with Giemsa
stain.
The bands
are very
large
regions
(-10?
bp)
that can
be used
to map chro-
mosomal translocations
or other
large
changes
in structure.
Lampbrush chromosomes
of
amphibians
and
polytene
chromosomes
of
insects
have
unusually extended
structures,
with
packing
ratios < I 00. Polytene
chromosome
s of
D. melano
-
gaster
are divided
into
-5000
bands.
These bands
vary in size by an
order
of magnitude,
with
an
average
of
-25
kb. Transcriptionally
active
regions can be
visualized
in even
more
unfolded
("puffed")
structures,
in which
material
is
extruded from
the axis
of the
chromosome.
This
may resemble the
changes
that
occur
on a
smaller
scale when
a sequence
in euchromatin
is transcribed.
The centromeric
region
contains
the
kine-
tochore,
which
is responsible
for attaching
a
chromosome
to the
mitotic
spindle.
The cen-
tromere often
is surrounded
by heterochro-
matin. Centromeric
sequences
have
been
identified only
in
yeast
S.
cerevisiae,
where
they
consist of short
conserved
elements.
These
ele-
lnCNTS, CDE-I ANd
CDE-III,
biNd
CBFI
ANd thc
CBF3 complex,
respectively,
and
a long
A-T-
rich region called
CDE-II
binds
Cse4
to
form a
specialized
structure
in chromatin.
Another
group
of
proteins that
binds
to this
assembly
provides
the
connection
to microtubules.
Telomeres
make
the ends
of chromosomes
stable.
Almost all
known
telomeres
consist
of
multiple repeats
in which
one strand
has
the
Crossing-over occurs
when
\,/
mismatch repair is absent
n
I
Y
28.20
Summarv
753

general
sequence
C"(A,1T)*, where n>l
and
m
=
| to 4. The
other strand
,
Gn(T lA\*, has a
single
protruding
end that
provides
a template
for
addition
of individual
bases in defined
order. The
enzyme telomerase
is a ribonucle-
oprotein
whose RNA
component
provides
the
template
for synthesizing
the
G-rich strand.
This
overcomes
the
problem
of the inability
to replicate
at the
very end
of a duplex. The
telomere
stabilizes the
chromosome
end
because
the
overhanging single
strand
G"(TlA)m
displaces
its homolog
in earlier
repeating
units
in the telomere
to form a loop,
so there
are no free ends.
References
W
Viral
Genomes Are Packaged
into Their
Coats
Reviews
Black,
L. W.
(1989).
DNA
packaging
in
dsDNA bac-
teriophages.
Annu Rev. Immunol.4),
267-292.
Butler,
P. J.
(19991
. Self
-assembly
of tobacco
mosaic
virus: the role
of an intermediate
aggregate
in
generating
both
specificity and
speed. Philos Trans
R.
Soc.
Lond.
B Biol.
Sci. )54,
5)7-550.
I(lug,
A.
(1999).
The tobacco
mosaic
virus
particle:
structure
and assembly. Philos
Trans. R
Soc.
Lond
B Biol.9ci.354,
fil-y5.
Mindich,
L.
(2000).
Precise
packaging
of rhe rhree
genomic
segments
oI the double-stranded-
RNA
bacteriophage
phi6.
Mitobiol
Mol Biol
Rev.
63, 149-160.
Resea
rch
Caspar, D.
L. D. and
I(ug, A.
(1962).
Physical
prin-
ciples in
the construction
of regular
viruses.
Cold
Spring Harbor
Symp.
Quant
Biol.27,
1-2+.
de Beer,
T.,Fang,
J., Ortega,
M., Yang,
Q.,
Maes,
L
,
Duffy,
C.,
Berron,
N., Sippy,
J., Overduin,
M., Feiss,
M.,
and Catalano,
C. E.
(2002).
Insights
into
specific DNA recognition
during
the
assembly
of a viral
genome
packaging
machine.
Mol
Cell 9, 981-991.
Dube,
P., Tavares,
P.,Lurz,
R.,
and
van
Heel,
M.
(1991).
The
portal protein
of bacteriophage
SPPI: a DNA pump
with ll-fold
symmetry.
EMBO
J 12,
t303_rlo9.
Fraenkel-Conrat,
H. and
Williams, R.
C.
(I955).
Reconstitution
of active
tobacco mosaic virus
from
its inactive
protein
and nucleic
acid com-
ponents.
Proc Natl.
Acad
Sci. USA 41,
690-698.
Iiang,
Y. J., Aerne,
B. L.,
Smithers, L.,
Haddon,
C..
Ish-Horowicz,
D.,
and Lewis,
J.
(2000).
Norch
signaliing
and
the synchronization
of the
somite
segmentation
clock.
Nature
408,
475-479.
CHAPTER
28 Chromosomes
Zimmern, D.
(1977).
The nucleotide
sequence
at
the origin for
assembly on tobacco mosaic
virus RNA.
Cell 11,46)-482.
Zimmern,
D. and Butler, P.
J.
(1977).
The isolation
of tobacco mosaic
virus
RNA fragments
con-
taining the origin for
viral assembly.
Cell
ll,
455462.
The Bacterial
Genome
Is
a Nucteoid
Reviews
Brock, T. D.
(1988).
The
bacterial nucleus:
a his-
tory. Microbiol. Rev.
52, 397
-41
l.
Drlica, I(. and Rouviere-Yaniv
J.
(1987).
Histone-
like
proteins
of bacteria. Miuobiol.
Rev. 51,
30r-3t9.
The
BacteriaI
Genome
Is
Supercoiled
Review
Hatfield,
G. W. and Benham,
C. J.
(2002).
DNA
topology-mediated
control of
global gene
expression in Escherichia
coli Annu.
Rev.
Genet.36, 175-203.
Resea rc h
Pettijohn,
D. E. and Pfenninger,
O.
(1980).
Super-
coils in
prokaryotic
DNA restrain
ed in vitro.
Proc. Natl. Acad.
Sci. USA 77
,
lj3l-1j35
.
Postow, L.,
Hardy,
C.
D., Arsuaga,
J., andCoz-
zarelli,
N.
R.
(2004).
Topological
domain
structure of
t}:'e
Escherichia
coli
chromosome.
Genes Dev. 18,
17 66-1779.
Chromosomes Have
Bandinq Patterns
Research
International
Human
Genome Sequencing
Con-
sortium.
(2001).
Initial
sequencing
and analy-
sis
of the human
genome.
Nature 409,
860-92t.
Saccone,
S., De Sario, A.,
Wiegant,
J., Raap, A. I(.,
Della Valle,
G., and Bernardi,
G.
(1993).
Cor-
relations
between isochores
and
chromosomal
bands in
the human
genome.
Proc.
Natl. Acad.
Sci. USA 90, 11929-ll9j3.
Venter,
J. C. et al.
(2001
).
The
sequence
of the
human
genome.
Science 291,
I 304-1
3 50.
The Eukaryotic
Chromosome
Is
a Segregation
Device
Review
Hyman,
A. A. and
Sorger, P. K.
(1995).
Srructure
and function
of kinetochores
in
budding
yeast.
Annu.
Rev.
Cell
Dev.
Biol 71,
47 l-495.
Centromeres
May
Contain
Repetitive
DNA
Review
Wiens,
G.
R.
and Sorger, P.
K.
(1998).
Centromeric
chromatin
and epigenetic
effects
in kineto-
chore
assembly.
CeIl 9), )13-316.
754

Research
Copenhaver, G. P. et al
(19991.
Genetic definition
and sequence analysis of Arabidopsis
cen-
tromeres. Science 286, 2468-247
4.
Haaf, T., Warburton, P.
E., and Willard, H. F.
(1992\
.Integration
of human alpha-satellite
DNA into simian chromosomes:
centromere
protein
binding and disruption
of normal
chromosome segregation.
Cell 7 O, 681-696.
Sun,
X.,
Wahlstrom, J., and I(arpen,
G.
\19971.
Molecular
structure of a functional Drosophila
centromere. Cell
9 l,
1007-10l 9.
Centromeres Have
Short
DNA
Sequences
in
S. cerevisiae
Reviews
Blackburn, E.
H.
and Szostak,
J. W.
(198a).
The
molecular structure of
centromeres and
telomeres. Annu. Rey- Biochem.
f
,
l8-194.
Clarke, L. and Carbon,
J.
(1985).
The
structure and
function of
yeast
centromeres. Annu. Rev.
Genet
19,29-56.
Resea
rch
Fitzgerald-Hayes,
M., Clarke, L., and
Carbon, J.
(1982lr.
Nucleotide sequence
comparisons and
functional analysis
of
yeast
centromere DNAs.
Cell 29, 2)5-244.
The Centromere Binds
a
Protein
Comolex
Review
Kitagawa,
I(.
and
Hieter,
P.
(2001).
Evolutionary
conservation between budding
yeast
and
human kinetochores.
Nat
Rev. Mol.
Cell
Biol.2,
678-687.
Resea
rc h
Lechner, J. and Carbon, J.
(
199 I
).
A 240 kd multi-
subunit
protein
complex,
CBF3,
is
a
major
component of the budding
yeast
centromere.
Cell 64,7
17-725.
Meluh, P. B. and I(oshland, D.
(1997).
Budding
yeast
centromere composition
and assembly
as revealed by in vitro cross-iinking.
Genes
Dev.
rt,340r-j4t2.
Meluh,
P. B.
et al.
(1998).
Cse4p
is
a component of
the core centromere of S. cerevisiae. Cell 94.
607-6t3.
Orliz, J., Stemmann, O.,
Rank,
S., and
Lechner,
J.
(1999).
A
putative protein
complex consisting
of Ctf 19, Mcm2 l, and Okpl represents a miss-
ing link in the budding
yeast
kinetochore.
Genes
Dev. 13, I 140-1 155.
Tetomeres Have
Simple Repeating
Sequences
Reviews
Blackburn, E. H. and Szostak, J. W.
(1984).
The
molecular structure of centromeres and
telomeres.
Annu Rev. Biochem
5).
163-194.
Zakian, V. A.
(
I 989
)
. Structure
and function
of
telomeres.
Annu
Rev. Genet.23,
579-604.
Research
Wellinger, R. J., Ethier,
I(., Labrecque,
P., and
Zakian, V. A.
(
I 996
)
. Evidence
for a new step
in telomere
maintenance.
Cell 85,
423-433.
Tetomeres Sea[
the Chromosome
Ends
Resea rc
h
Griffith, J. D. et al.
(19991.
Mammalian
telomeres
end in a
large duplex
l,oop. Cell 97,
5Oi-514.
Henderson, E., Hardin,
C. H., Walk,
S. K.,
Tinoco, I., and Blackburn,
E. H.
(1987).
Telomeric oligonucleotides
form
novel
intramolecular structures
containing
guanine-
guanine
base
pairs.
Cell 51,899-908.
I(arlseder, J
,
Broccoli,
D., Dai, Y.,
Hardy, S., and
de Lange,
T.
(I999).
p53-
and
ATM-dependent
apoptosis
induced by telomeres
lacking TRF2.
Science 28),
lJ2l-),)2r.
Parkinson, G. N., Lee,
M. P., and
Neidle, S.
(2002)
Crystal
structure
of
parallel quadruplexes
from human telomeric
DNA. Nature
417,
876-880.
van Steensel,
B., Smogorzewska,
A., and
de
Lange, T.
(1998). TRF2
protects
human
telo-
meres f rom end-to-end
f u sions. Ce
ll 92, 40
| 4 | 3.
Williamson, J.
R.,
Raghuraman,
I(. R., and
Cech,
T. R.
(1989).
Monovalent
cation-
induced structure
of
telomeric
DNA: the
G-quarter
model. Cell
J9,871-880.
Telomeres
Are Svnthesized
by a
Ribonucteoprotein
Enzyme
Reviews
Blackburn, E. H.
(
1 99 1
)
. Structure
and
function
of
telomeres.
Nature
)50, 569-573.
Blackburn,
E. H.
(1992).
Telomerases.
Annu. Rev.
Biochem 61,
ll3-129.
Collins,
K.
(1999).
Ciliate
telomerase
biochemistry.
Annu Rev. Biochem.
68,
187-218.
Smogorzewska,
A
and de
Lange,
T.
(2004). Regu-
lation of telomerase
by
telomeric
proteins.
Annu Rev. Biochem.T),
177-208.
Zakian, V.
A.
(1995).
Telomeres:
beginning
to
understand
the end.
Science
270,
l60l-1607.
Zakian, V. A.
(
I
996)
. Structure,
function, and
replication of.S
cerevisiae
telomeres.
Annu
Rev.
Genet.30,
l4l-172.
Resea
rch
Greider,
C. and
Blackburn,
E.
H.
(1987).
The
telomere
terminal
transferase
of
Tetrahyrnena
is a ribonucleoprotein
enzyme
with
two kinds
of
primer
specificity.
Cell 51,887-898.
Murray, A., and Szostak,
J. W.
(1983).
Construc-
tion of artificial
chromosomes
in
yeast.
Nature
305,
r89-r9).
Pennock, E.,
Buckley,
K., and
Lundblad,
V.
(2001).
Cdcl3
deiivers
separate
complexes
to the
References
755

telomere for end
protection
and replication.
Cell
104,387-396.
Shippen-Lentz,
D. and Blackburn,
E. H.
(1990).
Functional
evidence for
an RNA template in
telomerase.
Science
247,
546-552.
Teixeira,
M. T., Arneric, M.,
Sperisen, P.,
and
Lingner,
J.
(2OO4l
. Telomere
length homeo-
stasis is
achieved via
a switch between
telomerase- extendible
and
-nonextendible
states.
Cell
ll7,
)2)-)35.
@
Telomeres Are Essentia[
for
Survival
Resea rc h
Hackett,
J. A., Feldser, D. M.,
and Greider,
C. W
(2001).
Telomere
dysfunction
increases muta-
tion
rate
and
genomic
instability.
Cell lO6,
275-286.
Nakamura, T. M., Cooper, J. P., and
Cech, T. R.
(1998).
TWo
modes of survival
o{ fission
yeast
without telomerase.
Science 282, 493496.
Nakamura, T. M., Morin,
G.
B.,
Chapman, I(. B.,
Weinrich, S. L., Andrews,
W. H., Lingner,
J.,
Harley,
C. B., and Cech, T. R.
(1997).
Telom-
erase catalytic subunit homologs
from fission
yeast
and human.
Science 277, 955-959.
Rizki,
A. and Lundblad,
V.
(2001).
Defects in mis-
match repair
promote
telomerase-indepen-
dent
proliferation.
Nature 4ll
,
7l
j-7
16.
756 CHAPTER
28
Chromosomes

Nucleosome
CHAPTER OUTLINE
Introduction
The Nucleosome Is
the Subunit of
A[[
Chromatin
o
MicrococcaI nuclease
releases individuaI nucleosomes
from
chromatin as
115
oartictes.
o
A
nucteosome
contains
-200
bp of DNA, two copies of each
core
histone
(H2A,
HZB, H3, and H4).
.
DNA is wrapped around
the outside
surface of the
protein
0cramer.
DNA Is Coiled in Arrays
of
Nucleosomes
.
>95%
of the
DNA is recovered in nucleosomes
or
multimers
when micrococcal nuclease
cleaves
DNA
of chromatin.
r
The
length of DNA
per
nucteosome varies for individuat tis-
sues in a range from 154 to 260 bp.
Nucleosomes Have
a Common Structure
.
NucteosomaI
DNA is divided into the core
DNA
and [inker
DNA
depending on
its
susceptibitity to
micrococca[
nuctease.
.
The
core
DNA is
the length of 146 bp that
is found
on
the
core
particles produced
by
prolonged
digestion with
micro-
coccaI nuctease.
o
Linker DNA is
the
region
of 8 to
114
bp that
is
susceptibte
to early cleavage
by the enzyme.
r
Changes
in
the length of [inker
DNA
account
for the varia-
tion
in total
length of
nucleosomal DNA.
.
H1
is
associated with linker DNA and
may
lie at the
point
where DNA enters and [eaves the nucleosome.
DNA Structure Varies
on
the Nucleosomal Surface
.
DNA
is wrapped
1.65
times around the
histone octamer.
.
The structure of the DNA is altered so that
it has
an
increased
number of base
pairs/turn
in
the
middl.e, but a
decreased
number
at the ends.
The Periodicity
of
DNA
Changes on
the Nucleosome
r
-0.6
negative turns of DNA are absorbed by the
change in
bp/turn from 10.5 in solution to an average of
10.2 on the
nucleosomaI
surface.
which
explains
the [inking-number
parao0x.
Organization
of the Histone 0ctamer
r
The
histone
octamer
has
a
kernel of an H3z-H4z tetramer
associated
with
two
H2A-H2B dimers.
.
Each
histone is
extensively
interdigitated with
its
partner.
.
A[[ core
histones have
the structural
motif of the
histone
fold. N-terminaI taits extend out of the
nucteosome.
The
Path of
Nucleosomes
in the Chromatin
Fiber
.
L0 nm chromat'in
fibers are
unfolded
from 30
nm fibers and
consjst
of a
string of
nucleosomes.
.
30
nm fibers
have s'ix
nucleosomes/turn,
which are
orga-
nized into a
sotenoid.
.
Histone H1
is required
for formation ofthe
30
nm fiber.
Reproduction
of
Chromatin
Requires
Assemb[y
of
Nucteosomes
r
Histone octamers
are
not conserved
during
replication, but
H2A-H2B
dimers and
H32-H42
tetramers are
conserved'
.
There are different
pathways for the assembty
of nucteo-
somes
during
replication
and
independentty
of reptication.
.
Accessory
proteins
are
required to assist
the assembly
of
nucte050mes.
.
CAF-1
is an assembty
protein
that
is Linked
to the PCNA sub-
unit
of the
rept'isome;
it is required
for deposition
of
H3
z-H4z
tetramers
foltowi
ng reptication.
r
A different assemb[y
protein
and
a variant
of histone
H3
may be used
for
reptication-independent
assembty.
Do Nucleosomes
Lie
at
Specific
Positions?
r
Nucteosomes
may form at specific
positions
as the
resutt
either
of the [oca[
structure
of DNA
or of
proteins
that
inter-
act
with specific
sequences.
o
The
most common
cause
of nucteosome
positioning
is when
proteins
binding
to
DNA establish
a boundary.
o
Positioning
may affect
which regions
of DNA are
in the
finker
and
which
face of DNA
is exposed
on the
nucleosome
su
rfa ce.
Are Transcribed
Genes
0rganized
in Nucleosomes?
.
Nucleosomes
are found
at the same
frequency
when
tran-
scribed
genes
or
nontranscribed
genes
are digested
with
micrococcaI
nuclease.
.
Some
heavily
transcribed
genes
appear
to be except'ionaI
cases that
are devoid
of nucteosomes.
Histone
0ctamers
Are
Disptaced
by
Transcription
o
RNA
potymerase disptaces
histone
octamers
during tran-
scription
in a
model system.
but
octamers
reassociate
with
DNA as
soon as the
potymerase has
passed.
r
Nucleosomes
are
reorganized
when
transcription
passes
through
a
gene.
Continued on
next
page
757

?DM
@
Nucteosome Displacement
and
Reassembly Require
SpeciaI Factors
r
Ancitlary
factors
are
required
both
for
RNA
potymerase
to disptace
octamers during
transcription
and for the histones
to
reassemb[e
into
nucleosomes
after
transcri
ption.
Insulators
Btock
the Actions
of
Enhancers
and Heterochromatin
Insutators
Can
Vary in
Strength
o
Insulators
can differ
in
how
effectively
they bl"ock
passage
of an activating
signat.
DNAase
Hypersensitive
Sites Reftect
Changes in
Chromatin Structure
.
Hypersensitive
sites are found
at the
pro-
moters
of expressed
genes.
o
They
are
generated
by the binding
of tran-
scription factors
that displace histone
octamers.
Domains Define
Regions That
Contain
Active
Genes
r
A domain
containing a transcribed
gene
is
defined by increased
sensitivity to
degra-
dation by DNAase I.
An LCR May
ControI
a
Domain
o
An
LCR is located
at the 5'end
ofthe domain
and consists of several hypersensitive
sites.
What
Constitutes
a Regutatory Domain?
.
A domain
may have an insulator.
an LCR.
a
matrix
attachment site.
and transcription
unit(s).
Summarv
?EfB
JEETI
Insutators
are
able to btock
passage
of any
activating
or inactivating
effects from
enhancers,
silencers,
and LCRs.
Insulators
may
provide
barriers
against the
spread of heterochromatin.
Insulators
Can
Define
a
Domain
.
Insulators
are speciatized
chromatin
struc-
tures
that
have
hypersensitive
sites. Two
insulators
can
protect
the
region
between
them from a[[
external effects.
Insutators
May Act in
One Direction
o
Some insutators
have
directionatity.
and
may
stop
passage
of effects in
one direc-
tion but not
the other.
@
Introduction
Chromatin
has a compact
organization
in which
most DNA
sequences
are
structurally inacces-
sible
and functionally
inactive.
Within
this mass
is the
minority
of active
sequences.
What is the
general
structure
of chromatin,
and what is
the difference
between active
and inactive
sequences?
The
high overall
packing
ratio
of
the
genetic
material
immediately
suggests
that
DNA
cannot
be directly
packaged
into
the final
structure
of chromatin. There
must
be hierarchies
of organization.
The
fundamental
subunit
of chromatin
has the
sqme type
of design in
all eukaryofes.
The nucleo-
some
contains
-200
bp
of
DNA,
organized
by
an
octamer
of small,
basic
proteins
into
a bead-
like
structure.
The
protein
components
are his-
tones.
They
form an interior
core; the DNA
lies
on the
surface
of the
particle.
Nucleosomes
are
an invariant
component
of euchromatin
and
heterochromatin
in
the interphase
nucleus
and
of mitotic
chromosomes.
The
nucleosome pro-
vides
the first
level
of organization, giving
a
packing
ratio
of
-6.
Its
components
and
struc-
ture are
well
characterized.
The
second
level
of organization
is the
coil-
ing
of the series
of nucleosomes
into a helical
il:l1JffiTi"[1':,',f
:i:ffi,tlil.'"'ff
if
#:
mitotic
chromosomes
lsee
Figure
28.11).In
chromatin
this brings
the
packing
ratio
of DNA
CHAPTER
29 Nucteosomes
to
-40.
The
structure
of this fiber requires
addi-
tional
proteins,
but is not
well defined.
The
final
packing
ratio
is determined
by the
third level of
organization, the
packaging
of the
30 nm fiber itself.
This
gives
an overall
packing
ratio of
-I000
in euchromatin,
cyclically
inter-
changeable
with
packing
into
mitotic
chromo-
somes to
achieve an overall
ratio
of
-10,000.
Heterochromatin generally
has a
packing
ratio
of
-10,000
in
both interphase
and
mitosis.
We need
to
work through
these levels
of
organization to
characterize
the events
involved
in cyclical
packaging,
replication,
and transcrip-
tion. We
assume that
association
with additional
proteins.
or modifications
of existing
chromo-
somal
proteins,
are involved
in
changing
the
structure
of chromatin.
We do
not know
the
individual
targets for
controlling
cyclical
pack-
aging. Both
replication
and transcription
require
unwinding
of
DNA,
and thus
must involve
an
unfolding
of the
structure
that allows
the rele-
vant
enzymes
to manipulate
the DNA.
This
is
likely
to involve
changes
in
all levels
of
organization.
When
chromatin is
replicated,
the
nucleo-
somes must
be reproduced
on both
daughter
duplex
molecules.
In addition
to
asking
how
the nucleosome
itself
is assembled,
we must
inquire
what happens
to
other
proteins
pres-
ent in
chromatin. Replication
disrupts
the struc-
ture
of chromatin,
which indicates
that it
both
poses
a
problem
for
maintaining
regions
with
758
specific structure and
offers an opportunity to
change the structure.
The mass
of
chromatin
contains up to twice
as much
protein
as DNA. Approximately half
of the
protein
mass
is accounted for by the nucle-
osomes. The mass of RNA is < l0%
of
the mass
of DNA. Much of the RNA consists
of
nascent
transcripts still associated with the template
DNA.
The nonhistones include all
the
proteins
of chromatin except the histones. They are more
variable between tissues and
species,
and they
comprise
a smaller
proportion
of the
mass
than
the histones. They also comprise a much larger
number of
proteins,
so that any individual
pro-
tein is
present
in
amounts much smaller than
any
histone.
The functions
of
nonhistone
proteins
include control of
gene
expression and
higher-
order structure. Thus RNA
polymerase
may be
considered
to be a
prominent
nonhistone. The
HMG
(high-mobility group) proteins
comprise
a discrete and well-defined subclass of
nonhis-
tones
(at
least some of which are transcription
factors). A major
problem
in
working
with other
nonhistones
is
that they tend to be contami-
nated with other nuclear
proteins,
and so far it
has
proved
difficult to obtain those nonhistone
proteins
responsible for higher-order structures.
The Nucleosome Is
the
Subunit of
Atl.
Chromatin
r
MicrococcaI nuclease releases individual.
nucteosomes from chromatin as 11S
particles.
.
A
nucteosome
contains
-200
bp of
DNA, two
copies of each core
histone
(H24,
H2B. H3.
and H4).
e
DNA is wrapped around the outside surface of the
protein
octamer.
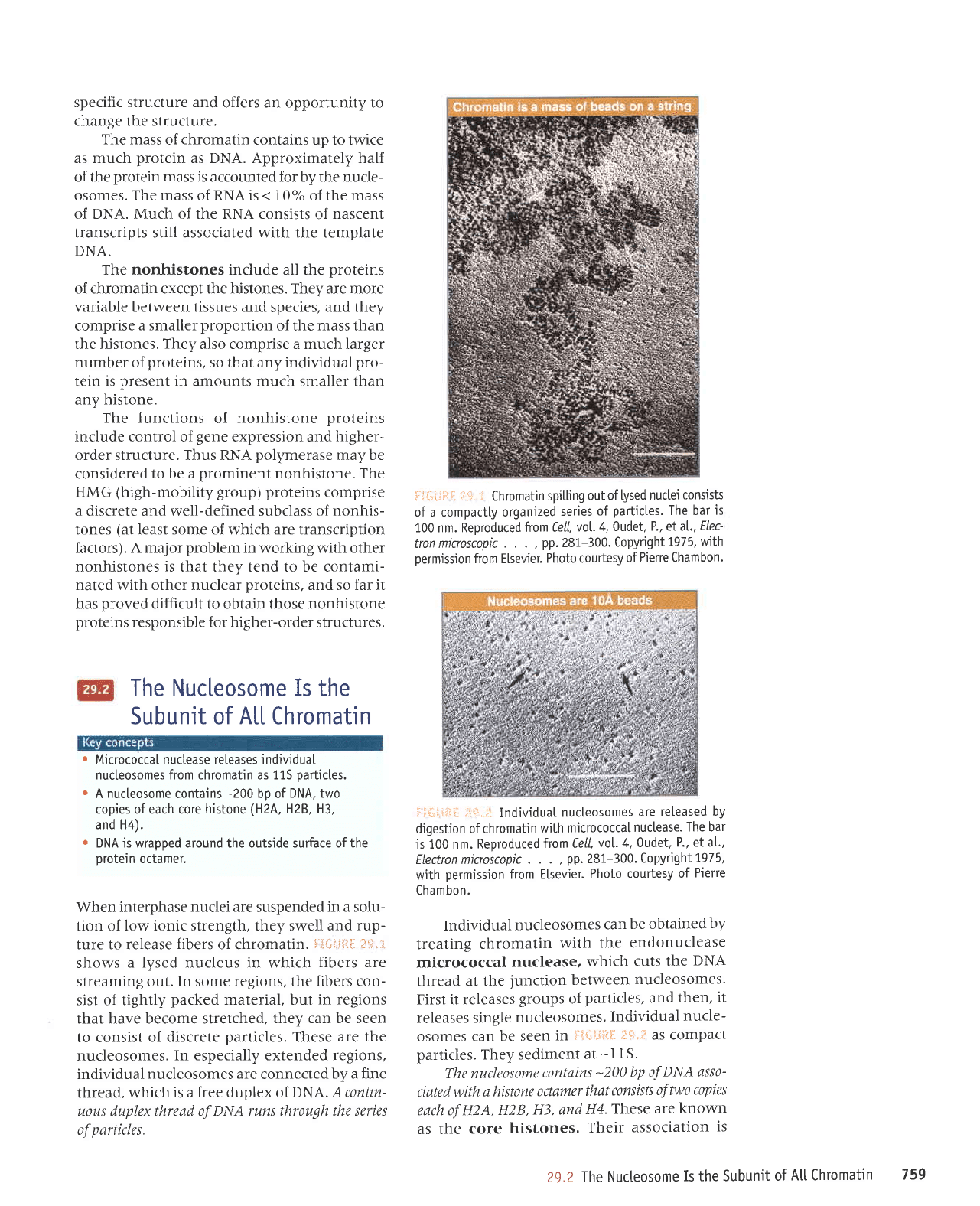
When interphase
nuclei are
suspended
in a solu-
tion of
low ionic strength, they swell and
rup-
ture to release
fibers
of chromatin.
$'i{;i"J{":*:
:li;.1,
shows
a lysed nucleus in which
fibers are
streaming
out. In some regions, the
fibers con-
sist of tightly
packed
material, but in
regions
that have become stretched,
they can be seen
to consist
of discrete
particles.
These
are the
nucleosomes.
In especially extended regions,
individual nucleosomes are connected
by a fine
thread, which
is a free duplex of DNA. A contin-
uous duplex thread of
DNA runs
through
the series
of
particles.
i:iri
iii'lL
iill.
"l
Chromatin
spitting
out of tysed
nuctei consists
of a compactly organized
series
of
particles.
The bar
is
100 nm. Reproduced
fron Cell,
vot.
4, Oudet.
P., etal.,
ELec-
tron microscopic
. . .
.
pp.
281-300.
Copyright
1975,
wlth
permission
from Elsevier.
Photo courtesy
of
Pierre Chambon.
Fiili,jiii:
j':;r,.,
IndividuaI
nuc[eosomes
are
reteased by
digestion of chromatin
wjth micrococcal
nuclease.
The bar
js
100 nm. Reproduced
from CelL,
vo[. 4,
Oudet.
P., et at.,
Electron
microscopic .
. .
,
pp.
281-300.
Copyright
1975,
with
permission from Etsevier.
Photo courtesy
of
Pierre
Chambon.
Individual
nucleosomes
can
be obtained
by
treating
chromatin
with
the endonuclease
micrococcal
nuclease,
which
cuts
the
DNA
thread at
the
junction
between
nucleosomes.
First it releases
groups
of
particles, and
then,
it
releases single
nucleosomes.
Individual
nucle-
osomes
can be seen
in
i,l{,i.iSil
;1F"f
as compact
particles. They sediment
at
-I
lS.
The nucleosome
contains
-200
bp of
DNA asso-
ciated with
a histone
octamer
that
consists
of two
copies
each of H2A,
H2B,
H3, and
H4.
These
are
known
as the
core
histones.
Their
association
is
29.2 Ihe
Nucteosome
Is
the
Subunit
of
Att Chromatin
759
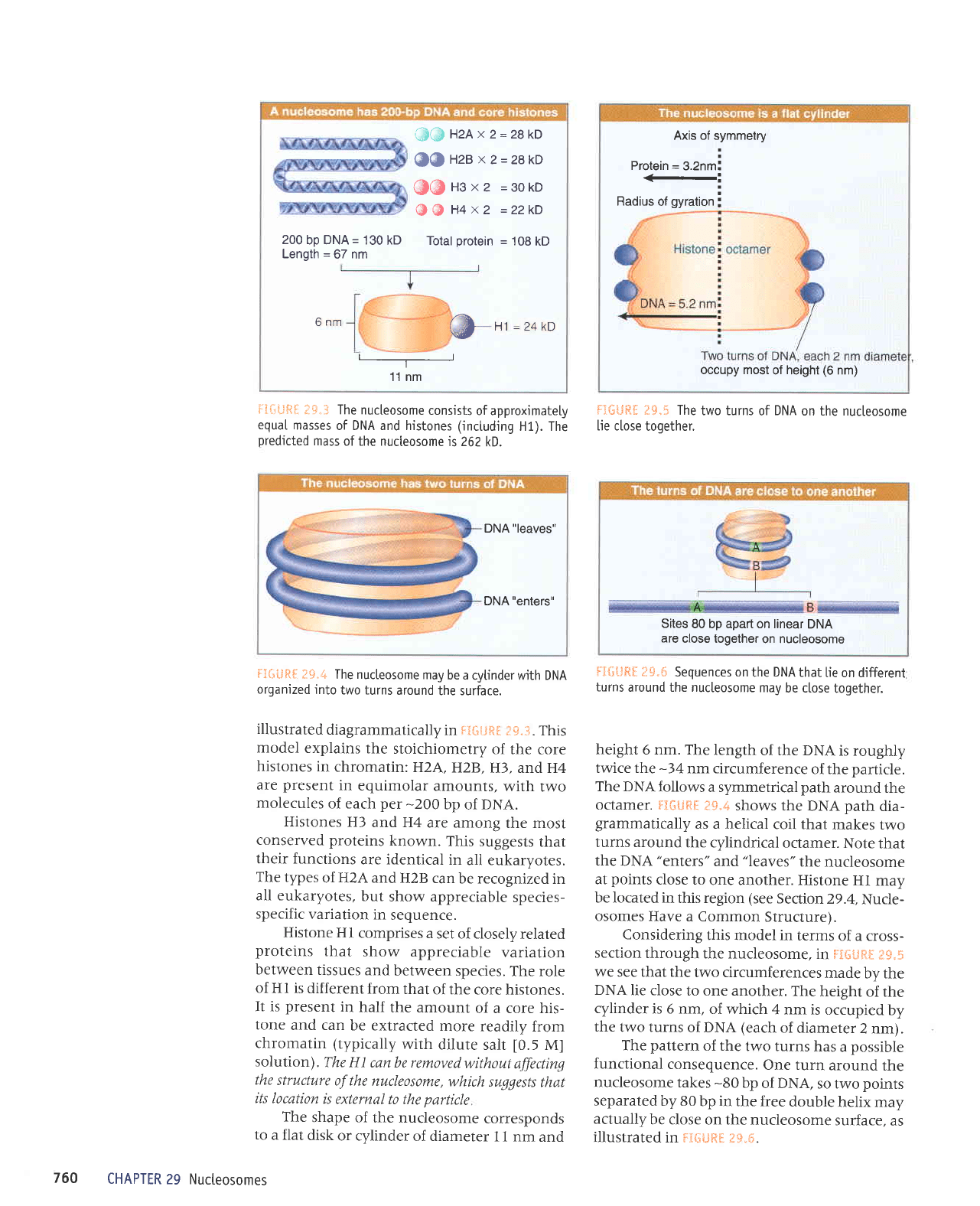
200
bp DNA= 130 kD
Length
=
67 nm
GG
H2A
x
2=z8kD
@@Hzex2=28kD
(lfi)ttsx2
=3okD
@ @
H4
x2
=22kD
Total
protein =
108
kD
6 nm
@
H1=z4kD
-___-------____-
11 nm
iliuRI
tF.3 The
nucteosome
consists
of approximately
equaI
masses
of DNA and histones (inctuding
H1). The
predicted
mass
of the nucleosome
is 262
kD.
DNA "leaves"
DNA
"enters"
FiStiRg
*9.4 The nucteosome
may
be a
cylinder with DNA
organized
into
two turns
around
the surface.
illustrated
diagrammatically
in f.gGiJRf
*S"i. This
model
explains
the
stoichiometry
of the core
histones
in
chromatirL;
H2A,
H2B,
H3, and H4
are
present
in
equimolar
amounts,
with
two
molecules
of each
per
-200
bp
of
DNA.
Histones
H3
and H4
are among
the most
conserved proteins
known.
This
suggests
that
their functions
are identical
in
all eukaryotes.
The types
of H2A and
H2B can
be recognized
in
all eukaryotes,
but
show appreciable
species-
specific
variation
in
sequence.
Histone
Hl
comprises
a
set of closely
related
proteins
that
show appreciable
variation
between
tissues
and
between
species.
The role
of Hl
is different
from
that
of the
core histones.
It is
present
in
half
the amount
of a core his-
tone
and can
be extracted
more readily
from
chromatin (typically
wirh
dilure
salr
[0.5
M]
solution).
The H1
canbe removedwithout
fficting
the
structure
of the
nucleosome,
which
suggests
that
its location
is external
to the
particle
The
shape of
the nucleosome
corresponds
to
a flat
disk
or cylinder
of diameter
11 nm
and
CHAPTER
29 Nucteosomes
Axis of symmetry
Protein
=
3.2nm!
<--!
Radius
of
gyration
!
occupy most
of height
(6
nm)
ruSeiffE IS.5 The
two turns
of
DNA
on the nucleosome
lie ctose
together.
Sites
80 bp apart on linear
DNA
are
close together
on nucleosome
tlfi{JBfi
t9-$
Sequences on
the
DNA
that
lie on djfferent
turns
around the nucleosome
mav
be ctose
toqether.
height
6 nm.
The length
of the DNA
is roughly
twice the
-34nm
circumference
of the
particle.
The DNA
follows
a symmetrical
path
around
the
octamer. i:ISugE
t$.4
shows the
DNA
path
dia-
grammatically
as a helical
coil that
makes two
turns around
the
cylindrical
octamer.
Note that
the
DNA
"enters"
and
"leaves"
the
nucleosome
at
points
close to
one another.
Histone
Hl may
be located
in this region
(see
Section
29.4,
Nucle-
osomes Have
a Common
Structure).
Considering
this model in
terms
of a cross-
section
through
the nucleosome,
in
t'gfiti&fl
t$-S
we
see that the
two circumferences
made
by the
DNA
lie close
to one another.
The
height
of the
cylinder
is 6 nm,
of which
4 nm is
occupied
by
the
two turns
of DNA
(each
of
diameter
2 nm).
The
pattern
of the
two turns
has
a
possible
functional
consequence.
One turn
around
the
nucleosome
takes
-80
bp
of DNA,
so two
points
separated
by 80
bp
in
the free
double
helix may
actually
be close
on the nucleosome
surface,
as
illustrated
in $3SEJffifr
f S.{i.
760