Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.


Il9
sloll:l
uor.lPlr.idau
uolJ la^olau
ol rxstupqlaw
]upllodrul
ue sI uoqeulquolau
6'02
uorlef,rldJJ
eql uJr{J
'arnlJnJls
slr aJolseJ
ol
pJeMJoJ
>lJoJ
eql slloJ eselrleq e
'paJredeJ
uaJq seq
aSPuep eqt JJIJV
'JJnlJnJls
xJldnp e
rxJoJ 01 Jred spupJls
JJlqSnep Meu aql
pue
'pJeM
->lJeq
seloru.dlanrpayya
>goJ
aql uJqM
'rnJJo
ol
uollerSllu
qJuerq
s.&\ollp srql
'.{11erged
lseal
le
'salqruJssesp
snleredde uorterqdar aq1
'a8e
-urpp
Jql sJaluno)ue
ll
uer{M
pJeMJoJ
Surrroru
sdols >go;
Jr{I
'sfiets
>lJoJ
uorteJrldar
e uaqm
aruoJlno alqrssod
Juo sMoqs
S:'*i
.l5;j*ij
'etseluep
aql ssed,{q 01
lr
Molle
o1 ro
parredal
uJeq seq
a8e
-rupp
aql JJUE
perolsJJ
Jq
ot
>lJoJ
aqt
8ur^tro1p ut
pJAIoAur
are sLe,vrqled
rredar-uorleurqruoJJU
'a8eruep
Jo
etrs
eqt
lp
ilels
Leru >go; uorlerrlda.r
eql asneraq
'palerqdar
sI
VNO
pa8eruep
uaqazr
paluasard
sr uorlentrs
letrads
v'uorterrldarlsod
'sr
leqt
'aruanbas
pa8eruep
rqt
Jo
.ddor e
qrltr
xaldnp
puoJas
p
sr eJeql uaq.tr
Lpo
pasn
Jq upl
rredar-uorleurqurof,Jr
lnq
'Jrrrrl
.due
le
pJSn
eq aldnurrd ur ue)
sdeaaqted
rreda.r-uors
-rJX![
'uorlenqs
aqt
pue
a8eruep;o ad,h aqt
uodn
puadap
IIrM
pasn
sr le.trqted
qrlq6
'VN11
uI
a8euep rredar o1 sr(erraq1ed.,(ueru
arreq
slla)
IIV
'sese^losar
r\q
956
pue
xeldnp
e ol
pa1.ta^uor
eq Aeu
pue
uorpunfr\epqlog
e sP auPs aql sr
llo}
pellPls
eql
jo
e.ln]lnlls aLll
o
'prPMroJ
uo]
aql a^ou 01 asetrlaq P asn
puP
a6euep aq1
buuredar
Uelsar
fipu >lroJ
pa]le]s
!
r
'spuPlls
pazrsaqluAs
r{1nau om1
aql uaamlaq 6uured r\q esra,rer r{eu
IroJ
pellels
V
.
'VN0
ul
lttu
P ro alLs
pabeuep
p
slalunolue
1r
uaqM
11e1s
feu >1ro;
uorlerrldar
y
.
sjoill uor.lPlrldau
urorJ Ja^olau ol
usrueqlahl
luPlrodtul
uP sI
uoqeurquoleu
'(,{err,rqfed
tuereJJrp
e uorJ raqloup ,{q
palntttsqns
aq ueJ
uoIDunI Jruos Jprlord o1
pasn
Llensn
arudzua
uB
qllqM
ur ro) Surddepano.dleured
a:e
leql
sruals^s JredJJ alntrtsuo)
qllqM
'sJIll^IDe
raqlo
pue
'ase.rau,{1od 'asealJnu
Jo
>lJoM
-leu
e
purl
01
Dedxa
lsnru
J^A
'JrPdeJ-uorleu
-rqruo)eJ
ur [ruanr;;a
peJnpJJ
Moqs stuplnu
Jln esne)eq'luapuadapur Llarrlua
lou
are
sLe,,u
-rlled)alpue
J^n
JqI'peralur srle,raqted
aql
,uoq
ro,{e,raqted
qrea
o1 3uo1aq suorpunJ
r(ueru.troq
aurJJp
le^ louueJ
aM
'lurod
lueuodur
uP
sa>lelu
z(lulBlrarun
srqJ
'snJol
)aJ e se suorlrpuoJ
Jo
les
rJqloue uI
palelosl
uJeq eAPq o]
lno
suJnl
snJoll/'n P sP
pauPu
puE
suoulpuoJ
Jo
1as
Juo
ur
pJlelosr
uorl€lnru
e sJrurlJruos
lnq
'sluElnlx
aqt;o sadllouaqd
Jql uo
pJsPq
Jre SJUJB uoll
-eurquoJeJ
pue
Jreder
yo
suoueu8tsap aqJ
.puJ
/s
33JJ
e sarrnbar lum
ut uorlJeeJ aql teAJMoq
lsde8
le
uortf,unJ
o1
lqSnoqi
are
,{aq1
't(roltqtqut
st
qJIqM
'gSS
eqr
;o
aruasard aqt
;o
alrds ur a1q
-urasse
01 slueuplrJ eqt
roJ alqtssod
U
J>lptu 01
sr suorl)unJ
rrJql
Jo
euo
'VNCI
papuerls-afurs
uo
slueurelrJ
YJeu
Jo
uorleruJo; aql alourord
,{aq1
ggrag
pue
UOJaU
'xalduror;o
sad,{1 oazrl
rxJoJ
surJlord aq1
'A)aJ
pue
'O)aJ
E)aJ
:saua8
eeJqt
Jo
dno.r8 e suretuoJ,(emqted
{raU
aqJ
'uollJe
rteda.r
aql alalduor uJql
seIlIAIpe aseraruz(1od
pue
ase
-epnN
'g
1
'97
arnSrg
ur dals
prrautar
aql urrogad
ot
1r
s,lrolle spuprls
a18urs aSueqJXJ
01
VJaU
Jo
^tlllqe er{J
'VNO
papuerls-a13uls
p
qtl,lr
V)aU
Jo
uoltPr)osse
eql 01
peal
z(aq1
'(s,{em
lueJaJJIp
ur
q8noqtle)
VraU
Jo
uollre
Jq1 01 :otrd uo1l
-)unJ
qloq
s.,{.e^4nqled
dJeu
pue
)gJau
eqJ,
'JeIuIp
autptuur{d
e
lsed
SutlerUdar raqe
UJI
are
tpql
pueJls
ralq8nep e ut
sde8 aq1 8ur
-rredar
ur
pJAIoAur
sr
Lelrqled
dJeU
aqJ
'(srorrg
uorlerlldag
ruoJJ JaAoreU
01 uslueqJew
luel
-rodrul
ue sI uorteurquo)eg
'6'97
uorpag aas)
s>lJoJ
uorleJldar
pallets
Suruelsar
uI
pJAIoAUI
sr z(emqted
)gJJU
etfl'1ilt^
,ll suollf,unJ
lualeJ
-IIp
IIIJInJ
Laql
'pauqap
IIaM
os
tou
srpuu
pat
saAIoAur Jaqlo
eql
ipezuJlJPJeq)
IIaM
sI
puP
saua8
Sgtat
aql
sJAIoAur
ziea,rqted )_aJ euo
'(0E
se
Aueru se JIp
-upq
01 unrrJpeq
ad,{t-pp,rzr.
e
yo,{t1gqe
aqr
qllm
paredruor)
aruoua8 stl
uI sJalrrlp
Z
01
I<
JleJalol
louueJ
luPlnru
alqnop
aqt
r(q,u suteldxa
lI
'uop
-JunJ
VJaU
Jo
eJuasqe
eql
q
uopeJl1dar
o1 alt
-etsqo
1eqlel
e apnord
sJeIuIp
rql
lpq1
satTdurt
UnsJJ
srq1,
'sJalrrlp
aunuLqt
uJaMleq
aJuel
-srp
paDadxa
Jql
I{u,lr spuodsa::o;
azIS JSoqM
VNC
Io
sluaru8er;
arnpord slleJ -V)aJJ^n
ur
ygq
alerqdar ot
sldurallv
'saIlITIJeJ
Lrarrorar
pue
rredar
Surutetuar
Jql
IIe
saqstloqe
[11et1
-uJssa
aua?
ycat
aql
q
uollelnlu
lredar uors
-rJXJ
ur
tuJnrJJptor'g
ul'(E
l'61-€1
'61
sarn8rg
aas) saua8
ra"r aqr
dq
pagpuapl sI
to,
'g
ur rtedar
-uorleulquolJJ
JoJ
.{e,rzrqled
pdtrurrd
aq1
'(uatsz(s
rredar
uoISIf,xJ
ue ,{q
paaoruar
sr
eBeurep
Jql
illun
pue
ssJlun
al:zb uorlertldar
.{rana rat;e
paleadat
aq
lsnru
sluena rtedar
-uorleurquoJJJ
alrrps
aqt
q3noqrle)
uottrol
-sp
pur8tro
Jql
01
paulJuor
sr a8eruep
aql snqJ
'xaldnp
lelrrJou
e SurlerauaB
'trenv
Iensn
eq1
ur srsaqlu^ds
rredar
^dq
pallti
aq uer deE
aqr
lde8
e Sune;
pueJts pluared
lpruJou
e seq
xaldnp
sualsns
rtedeu
OZ
UlIdVHl Zl9
DPXa
eql
1!\Ou>I
l,uop
eM
'JrpctaJ-uoppulqruoJar
z(q
panrsar
aq
ueJ $lJoJ uortp)rtdar
pa1p15
'alrs
srql
le
palpJJJ
sr
>lpJrq
pupJts-elqnop
rJqloup
'>lJru
p
t)eJ
ul
q
a8e
-rupp
eql
JI
'uorlrppp
uI'spuprls
fuetuarualduror
;o
rred Jaqlra
sanpalJ
asplloseJ
p
Jr
palpJa
-ua8
sr
learq
puprls-elqnop
v
'sesp^losar
ro;
la8
-JB1 p
1r
sJ>leu
slql'syN( xaldnp
omt uJJAtaq
uorlpurquorar
d.q
pateeJJ
uorlrunI z(epr11o11
p
sp
arups aql dilerluassa
sr >lroJ
pallpls
eql
Jo
alnlJnJls eql
leql
sMoqs
0z'02
IungIJ
'vJau
Jo
'spua
,€
alPl
-rpur
sppeqmorJv
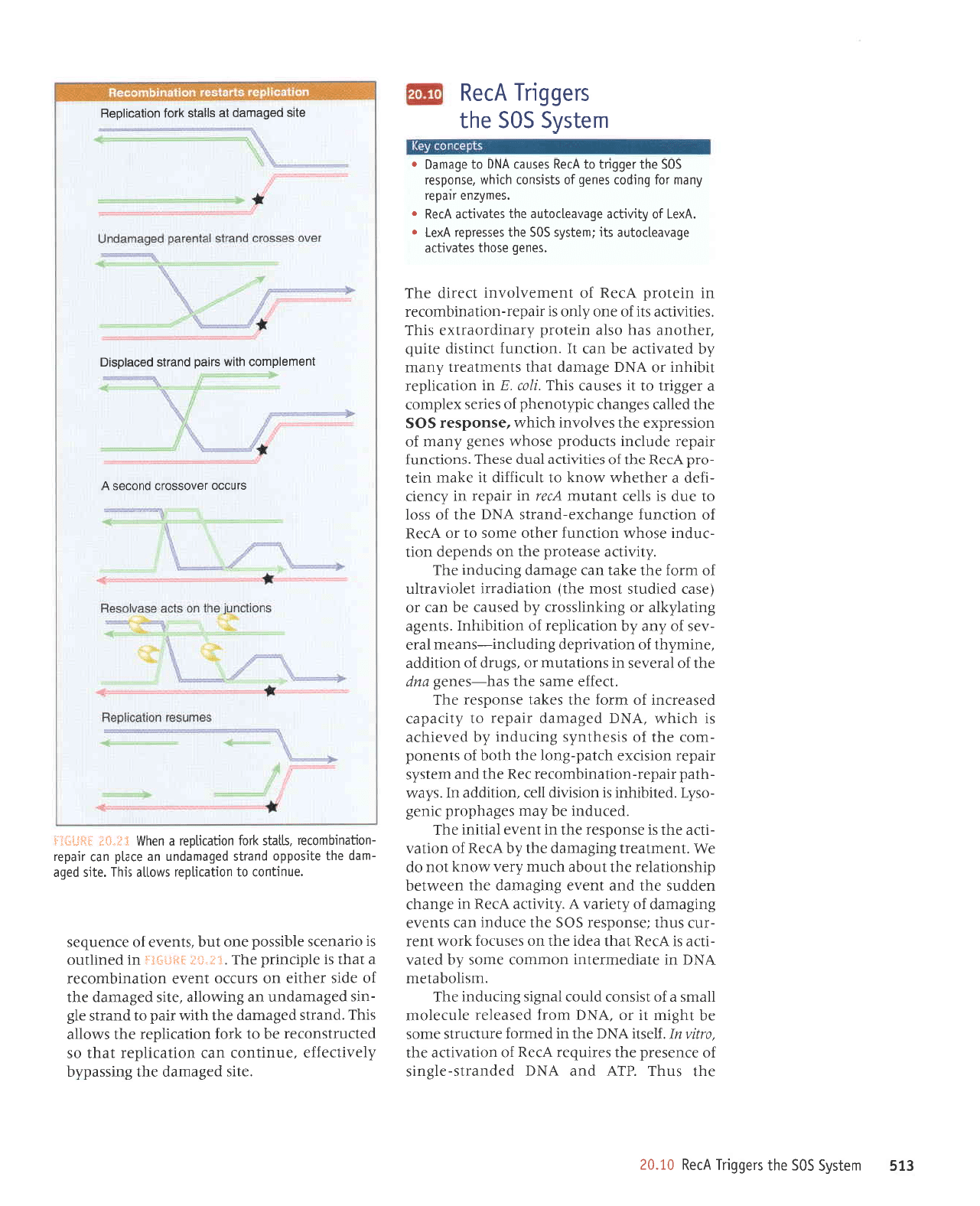
'ltru
p
surpluol
1r;t
abeuep
;o
a1s aql
lp
palerauab
sr
ES6
puolas
p
smoqs
Z llnsau
'uorpunl
aq1
1e
spuerls;o rred e
6uq1nr [q
paleraua6
st
leelq
pue.l]s-alqnop
e
lpql
smoqs
t llnsau
'llru
e surpluor aEeuep
jo
alrs aql
laqlaqM u0
puadap
sllnsat
aql
'sese^losal
r{q
r{ervr aues
oql ur
pe^losal
aq upl
pue
uorpun[ Aeprtlog
e salquesel
uo;
uorlerrldal
pallpls
p
Jo
atnllnlls
aqt
OZ.OZ
lungll
suorlJpJJ a8ueqrxa-puerls aq1 z(lqrssod-rrcdar
-uopeulqurof,er
asn deru
l(euqted
raqtouy
'erunsal
uer uorlerqdar
'parrcdar
uaaq spq
a8eruep Jr{l rJl;v
'aSeurep
er{l
Jo
JrpdaJ-uors
-rJXJ
ur
pallolur
sr
)gJaU
'luala
Euupts aql
sDetap
lpql
Josuas aql se 8upe,{1q1ssod
pue
>Fo;
uorlBrudar
pallpts
eql
lp 1l
01 Eutputq
r(q
VNq
papuerts-a18urs
azryqpls ot
V)aU
JoJ sr Lemqled
alqrssod aug
'(runrralJeq
Jql ut uoDJunJ roleur
Jraql Jq z(eru
slqt
'rre;
ur) uop)peJ slqt ur alor
tueuodrur
up alpq srualsls
)gJaU
pup
VJaU
eql'qlJ'g' u1
'seruzkua
rrBdar sarrnbar
{Jo}
uoll
-er{der
pellpls
e Sulpueq roy
z(euqred
aq;
'11I
aseraruz(lod
17NO
Iq
pareldar
ralel
sr
pup
'tJetsal
uoglerqdar aql JoJ
parnbar
sr
1
asererudlod
y1'.1q
'(uo4er11daU
ue$rg
01
paparN
sI eruosorurJd
aqJ
'/
I'8I
uorDas aas)
papelsar
sr uorlerqdal
pup
'Jlquasspar
uer snleredde
'spue,€
alplrpur
speaqmorJv
'aserrleq
e
fiq
pazAleler
uoqerbrur
qluplq-plpMroJ
r{q
parolser
sr
IroJ
aql
'parredar
uaaq seq
abeuep aql
lelJv
'rred
o1 spuerls ralqEnep
o/'^1 aql sMolle
uoJ
aq1 bursranax
'vNo
u] alrs
pabeuep
p
saqlear
1
ueqm
qlels
ryoJ
uoqerrldar
V
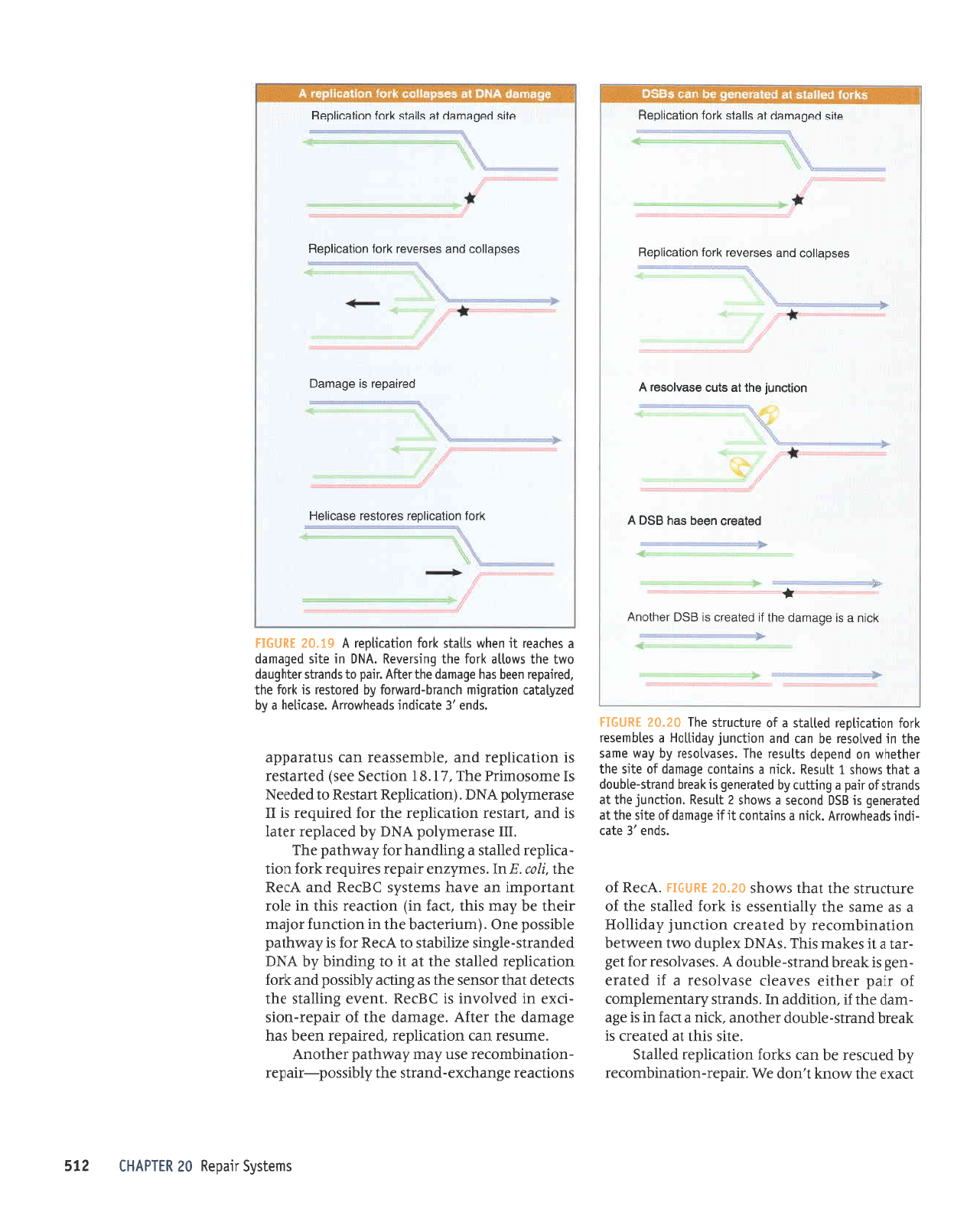
6I'02 lgngH
IJOI
uOrlecrloaj sarolsor asecrloH
perreder
sr e6eueg
sesde;;oc
pue
sesianar
1ro1
uonecrldeg
eys
pebeuep
le
sllets
1ro1
uorlecr;deg
Icru
e sr a6euep
oLll
Jl
polpoic
st
gSC
loqiouv
..+
pelearc
ueoq sEq
8so v
uqpun!
€ql
le
slm ese^losol
V
sesdelloc
puE
soslonol >1to1
uorleclldeg
elrs
pebeuep
le
sllels
lrol
uorlecrldeg
EI9
uets^s
sos
aqt sra66ul
vrau
0I.02
Jr{] snqJ
aIV
pue
vN(
prpuPrts-Jlburs
Jo
J)ueseJd Jql sarrnbJJ
vJJu
Jo
uorle^rlJe
eql
'0J1!^uI'Jlesll
VN(
Jql ur
peruJoJ
aJnDnrls Jruos
rq
lq8ru
1l
ro
'ygq
luorJ
peseslar
rlnralou
Ilerus
e
Jo
tsrsuof,
plnoJ
lpu8rs
Sunnpur aq1
'rusIoqelJru
vNC
ul JlerperurJlur
uoururoJ Jruos
^q
pele^
-llJP
sI
VIJU
leql
eJpr Jql uo
sesnJoJ
>lro1!{
lurr
-rn)
snql
lasuodsar
sos
Jql af,npur
upJ stue^J
Sur8euep
Jo
rllalren
V'^lrnrpe
V)eU
ur a8ueqr
uappns
Jql
pue
tua,ra
Sur8Burep Jqt
uJaMtJq
drqsuorlelar
Jql
lnoqe
qrnu
Lran ,lrou>l
tou
op
JM
'luauleJrl
Sur8euep
eqt ^q
vJJU
Jo
uorlp^
-rpe
aqt
sr asuodsar Jql ul
tuJ^J
Iprlrur
JqJ
'pJJnpu
aq,{eru saBeqdord
rrua8
-osA1
'patlqlqur
sr
uorsrlrp
IIJ)
'uourppe
u1
's.de,vr
-qled
rredar-uorleurquoraJ
JJU Jr{l
pup
uralsLs
rredar
uorsrJxa
qrled-3uo1
Jqt
qtoq;o
stuauod
-ruoJ
Jr{l
;o
srsaqlu.{.s Sunnpur Lq
pa.tarqre
sI
qJrqM
'ypq
pa8euep
rredar ot
.,(tnedBr
pJSeJJ)ur
Jo
urJoJ
JLll sJ{et asuodser
aq1
'DaJJJ
Jues Jql seq-sauaB
aup
Jqt
Jo lereles
ur suortptnu
ro
's8nrp
Jo
uourppe
'auruzi.qt
Jo
uorle,trrdap
8urpn1:ur-sueau
IeJa
-Aas
Jo
,(ue
Aq uorlerrldar
Jo
uortrqrqul
'slua8e
3urre1.{4e ro
3urluqsson ,{q
pasner
eq upr
Jo
(ase) perpnls
lsolu
aql)
uorterperrr
talor^prtln
Jo
ruJoJ aql e>lel
uer a8eruep Sunnpur
aq1
',{ttlrlre
aseatord
Jql uo
spuadap
uotl
-Jnpur
esor{^^ uorltunJ
raqlo auos 01
Jo
YJeu
Jo
uorpunl
aSueqrxa-pupJls
vNo
3ql
Jo
ssol
ol anp sr sllJJ
luelnru
V)dt
ur rrpdar
ur
,{luatr
-lJep
e JJqlJqM MOu>I
01
llnJrJJrp 1r
J>leIrI
ural
-ord
yrag
Jql
Jo
serlrlr1re
Ienp
aseqJ
'suortrunJ
rtedar
epnlJur spnpord
JSoqM saua8
Lueru;o
uorssa-rdxa
Jqt seAIoAur
qJIqM'asuodsar
56t5
Jq1
pJIIeJ
sa8ueqr
rlddtouaqd
yo
sJrrJS
xalduor
e ra33u1
ol
lr
sJSneJ srqJ
'qoJ 'E
ur uorlerrldar
tlqlqul
Jo
VNo
a8eruep
teqt
stuJrutean
r(ueru
Lq palenrpe
Jq
ueJ
lI
'uortJunl
purtsrp
alrnb
'rJqloue
seq osle urato-rd ,{reurproenxJ
sIqI
'sarlrnrDe
slr
Jo
euo ,{1uo
sr rredar-uorleurqruo)eJ
ur uratord
VJaU
Jo
tuaurJnlonul lJJJrp
JqI
'sauab
asoql
sale^qle
abenealrolne s1L 1uLa1s[s
S0S
aql sassaldar
yxal
o
'VXel
Jo
AlnLlre
abenealrolne oql sale^qle
!lo!
o
'saur{zua
tLpda.t
r\ueu io; 6uLpor saueD
Jo
slsrsuol
qrLr.ln
'asuodsar
S0S
aql ra66u1 o1
VlaU
sasnpl
y116
o1 abeueg
o
ualsns
sOs
aql
sra66u1
VlaU
'atrs
pa8euep
aqt Sutssed,{q
LlanrDalya
'JnurtuoJ
upJ uolte)Ildar
1eq1
os
pJlfnJtsuo)Jr
Jq ol
IloI
uorlpJlldar
aqt sMolle
srqJ
'puens
pa8euep
aql
qtprzr
rred
o1
puerls
a13
-urs
pa8eruepun
up Sulraolp
'a1rs
pa8eutep
aql
JO
JprS JJr{lre
uo srnJJo
IuJAJ
uoIlPUIqlUO)eJ
e
teqt
sr aldourrd
JVJ
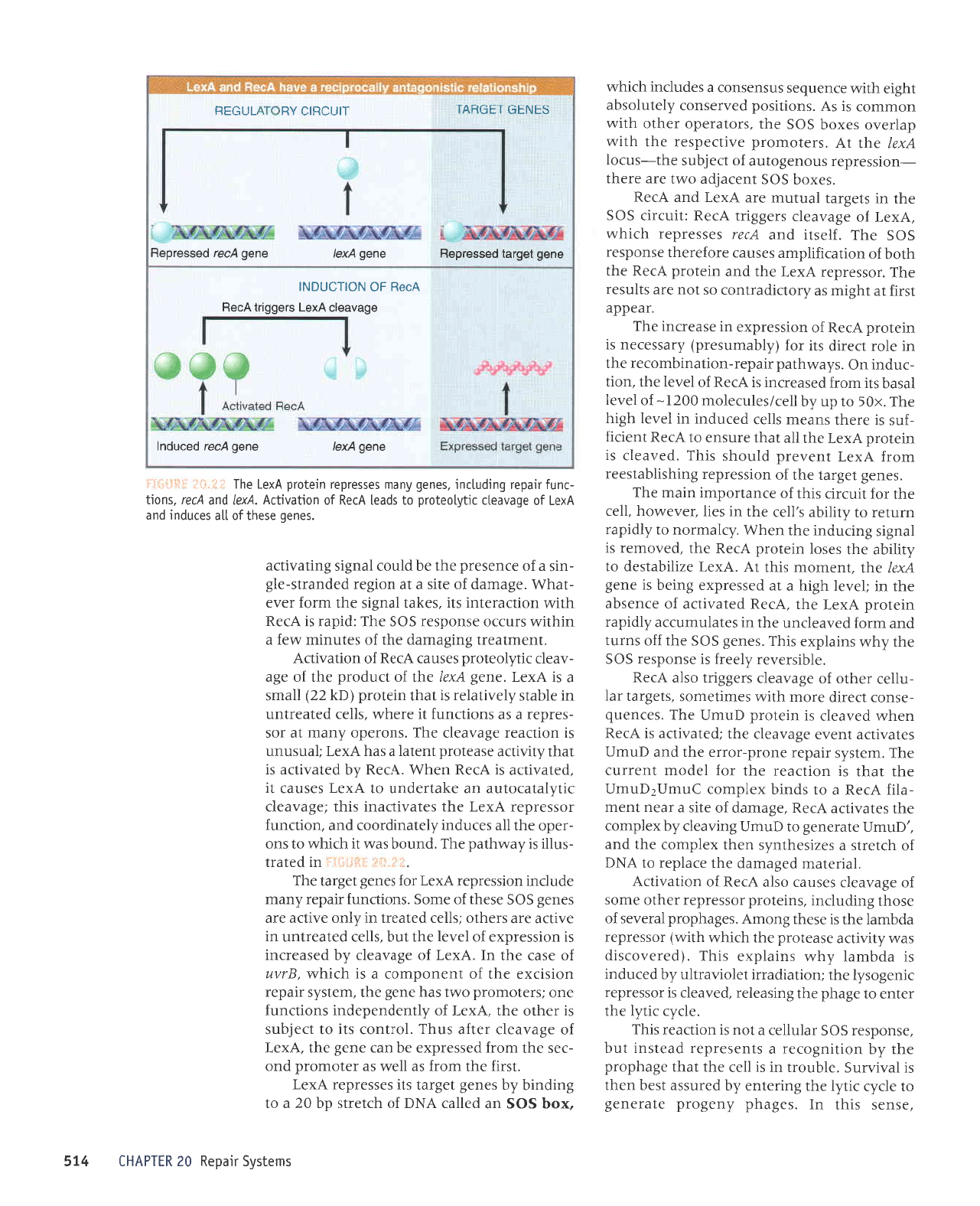
'l"i'*c1
tst't*l:i
uI
pJulltno
sr orJpue)s
alqrssod euo
lnq'sluJ^a;o
a;uanbas
'enurlu0)
01 uorlPltl00r
s^40lle stqf
'olts
paDe
-upp
aqt alLsoddo
puerls
pa6euepun
ue areld uer rledet
-uo4Purquolar
'sllPls
)loj
uoqeltldol
P uaqM
i,I'{ii:
!"sll':}i:i
srncco
ro^ossors
puocos
v
lueueldu.roc
qlnr
sred
pueJls paceldsl6
elrs
peoeuep
le
sllEls
tlrol uotleotldeH
'JSUJS
srql uI
'sJBpqd
.r{.ua3o.rd
ateraua8
ot alrrb rr1l.1 aqf
Surralua Lq
parnsse
tsaq
uJql
sr
IeATAJnS
'Jlqnon
ur
sr
IIat
Jr{t
tpqt
a8eqdord
Jql
^q
uorlruSorar
e stuasardal
ppelsur
tnq
'JSuodsJJ
SOS
relnlleJ
p
tou
sr uollJpJJ
slqJ
'3ll^)
JrlAI Jql
rJlue 01 a8eqd aql
Sursealar
'pa^ealJ
sr rossardar
rruaSosLl aqt
luorterpeJJr
tJlorleJtln
Lq
parnpur
sr epqtuel .{qlt
sureldxJ
srqJ
'(paranorsrp
se,ra Lltntlle
eseJtoJd
aql
qJrqM
qlna)
rossardar
epqruel
aql sr Jsaql
Suouy'sa8eqdord prrnas
Jo
asoql Surpnput
'suralold
rossarda-r
JJqto aluos
;o
a8e,realr
sesneJ
osle
VJaU
Jo
uortpnrllv
'lpuatpur
pa8eruep
aql areldar
ot
VN(
Jo
qJlJns
e sazrsaqlu[s
uJql xaldruor
Jq]
pue
',qnrufl
atBraua8
ot
Onrun
Suneap
Lq xalduor
Jql SJlenrlJe
yraS
'aSeupp
Jo
Jlrs
e reJu
lueru
-elII
VJJU
e
ol spurq xalduror
)nuflzqnurn
Jql
leql
Sr uorl)eal
eql ro}
Iapou
luJJJnJ
aq1
'rualsAs
rredar
auord-rorra
eqt
pue
(nrun
salenrtf,e
tuela
eBeAeall
aql
jpJlelrlJe
sr
YJJU
uJr{M
pJ^eJ1:
sr
uralord
Cnrun
aq1
'saruanb
-esuoJ
l)Jrrp
Jrorx
qlrM
sJrurlJruos
's1a3rel
re1
-nllar
reqlo;o
a8enealr
sraSSut
osle
vJeU
'elqrsJJAJJ
,{.1aar;
sr asuodsar
595
aql .{q.Lr
sureldxa
srql
'saua8
SOS
Jqt
JJo
surnl
pue
ruro;
pJApJIJun
aqt ur
setEInLLrn)Je.{lprder
uralord
VXJ'I
Jr{t
'VJJU
pJteAIDe
Jo
JJuJsqe
Jqt ut l1a,ra1 q8rq
e
te
passardxa
Suraq
sr aua8
vxal
Jql'tuaruolu
srql
lv
'vxJf
ezrlqplsap
01
,{1r1qe
Jql sJsol
uratord
VJJU
Jqt
'penoruJl
sr
leu8rs
Sunnpur
aqt
ueq6
'Lrleurou
or r(lprder
uJntJJ ot ^llllqp
s,lle)
aqt
ur sJrl
'JelJ,lroq
'llJJ
Jql
rol
lrn)Jr)
srr.l
I
Jo
e:uplrodur
utpul JqJ
'saua8
laSret
aqt
;o
uorssardar
Surqsqqelsaar
ruorJ
VXaf
tuanard
plnoqs
srql
'paneall
sr
uratord
VXef
Jql
IIe
leql
Jrnsue
ol
vJaU
luarJrJ
-Jns
sI JJJqI
suearu
sllel
pJtnpu
ur
1a,ra1
q8rq
rqJ
'x0E
o1 dn z{q gar/salnrJlotu
00ZI-
}o la^al
IPSPq
s1r ruoJJ
peseJJJur
sr
v)eu
Jo IJ^JI
Jql
'uoll
-Jnpur
u6'sLe.Lrqled
ledar-uorleurqruoJJJ
Jql
ur eloJ
tJaJrp
str roy
(l.lqeunsard)
Lressarau
sr
utalord
yrag;o
uorssardxe
ur asperf,ur
eqJ
'readde
lsJrJ lp lqStu
se ^,fuolrrperluoJ
os
lou
aJe sllnsJr
aql
'rossardJJ
vxJf
eqt
pue
uratord
VJaU
Jql
qloq
Jo
uotlerqlldrue
sJSneJ
JJo]eJeqt
asuodsar
SOS
aql
'JIJstr
pue
y;a;
sassardJt qJrqM
'VXJ'I
;o
a8e,teap
sraSSrrl
VJeU
:trnJrlJ
SOS
aql
uI sla8rel
lentnru
aJp
VXef
pue
V)aU
.SAXOq
SOS
IUJJEtPP
OMl
JJP JJJqI
-uorssaldar
snouaSolne
;o
palqns
eql-snJol
VXal
ertrt
tv
'srJtouord
a,trtradsar
Jql
qlr,tr
deparro
sJxoq
SOS
aqt
'sJoterado
raqto qtr,u
uoruruor
sr sV
'suorlrsod
pa,rrasuor
,{1a1n1osqe
IqBIJ
r{lrm aruanbas
snsuesuoJ
e sepnlJur qJlq&r
suralsr{5 rredag
67
UlldVHl ,19
'xoq
SOS
ue
pallpr
vN11
Jo
qrtrrls
dq
0Z
e ol
Surpurq Lq saua8
tJBJpt
slr sassa:dar
yxal
'tsJIJ
Jql uoJJ sp
IIJM
se raloruord
puo
-lJS
Jql uor;
passardxJ
Jq ueJ aua8 aqt
'Vxa'I
;o
a8enealr
rJUe snql
'loJluoJ
slr ol
tralqns
sr rer{lo Jqt
'VXJ'I;o
.{.lluapuadJpur
suorlJunJ
euo
lsrJloruord
o.r,rl seq aua8 aq1
'ruals.ds
rredar
uorsrf,xe
aql
Jo
luauodruoJ
e sr rtrJrrtrlv'
'gJAn
Jo
Jspf, aql
q
'VXef
Jo
aBe,Leap Lq
paseanur
sr uorssardxa
Jo IJ^JI
Jql
lnq
'sllJJ
peleJrlun
ur
JAI])E EJE SIJqIO ISIIJ)
PJTPJJ]
UI
.dlUO
JAI}JP JJE
saua3
5gg
eseqt
Jo
euros
'suortJuny
rredar.{ueru
epnpur uorssardar
VXJ1
JoI saua8
ta8ret
aq1
'i,l'r:r,
:iiii;::i:l
uI
PeleIl
-snllr
sr
^de.trqted
eqJ'punoq spM
tr
qlrqu
ot
suo
-rado
aqt
1e
sarnpur.{latpurpJooJ
pue
'uorlJunJ
rossa:dar
vxJf
Jql sJlelrlJpur srql
la8enealt
rr1,{1e1ero1np
ue J>leuapun ol
VXJf
sasnp)
1r
'pJlPArllP
sr
vJeu
uaqM
'y:ag
,lq
pJlp^rlre
sr
leqt,(lurDe
aseatord
tuJtel
p
seq
VXa'I
{ensnun
sr uortf,pJJ a8e,tea1r
aq1
'suo:ado
,{.ueru
le
ros
-sardar
e se suorlJunJ
ll
JJJqM
'sllJr
palearlun
ur rlqpls,{1aar1e1ar
sr
lpql
ural0rd
((>I
77)
Ileus
e sr
VXJ'I
'aua8
yxal
JrIt
Jo
Dnpo:d
aql
;o
a8e
-neap
rrlj.10a10rd
sasner
\/Jeu
Jo
uorlP^rlJv
'lueruleaJl
Sur8eruep Jql
Jo
sJlnurru .,t,t.J]
p
ulqlrM sJnJJo asuodsar
SOS
JqJ
:prder
sr
yrag
qll,lr
uolDerelur
slr
'sa>1e1
leu8rs
JrIt ruJoJ rJ^J
-teq6
'a8eruep
Jo
Jlrs
p
tp
uor8al
papuerls-a13
-urs
p
Jo
aruasard aql eq
plno)
leu8rs
3ur1e.a,r1re
'saue6
aseql
jo
lle
salnpur
pue
VXol
Jo
ebeneelr
rLy{1oa1ord ol sppal
VraU
Jo
uoqe^rllv
'Vxal
pue
V)at'suotl
-run1
rredar 6uLpnlruL'seuab [ueuL
sasserder uralord
yxal
aqI
i:i]"i:r;i;
::iifll.iiJ
aue0yxel eueiycet
pocnpul
ebeneelc
yxel
sre66r.r1
ycag
VCOH JO NOIICNONI
aue6
1e6re1
pesserdag
eue6yxel
aue6ycet
pesseldeg
SSN3S
t3gHvt
J-lnCHlC AH ln93H

9r9
suels^S
riedau
pa^rasuol
a^eH
qla]
rqofiplnl
II'02
pallPJ
sJueE
lq3la
olur
IIPJ
suorlelnw
'stJnppe
.{4nq
raqro
pue
sJarurp aurpnurr.{d JSTJXa
tou
-upJ
sluJrled
4y
ruor; slsplqoJqrd
'rredar
uots
-oxa
ur.{:ualJlJep
p
z{.q
pasner
sr eseasp aqJ
'(spayap
eJJAJS aroru
sJrurlJruos
pue)
s;ap
-Josrp
uDIs ur stlnseJ Lruanqap
eql'tJlornerlln
'relnlttrBd
uI
pue
'lq8r1uns
o1 ,htnrlrsuas
-raddq
ur
3ur11nsar aspasrp elrsse)JJ e
'(aX)
runs
-otuaru8rd
eruJepoJJx
sr esaqt
Jo
pale8usa,rur
lsJq
JqJ'srapJosrp d;etrparaq upunq urcgar
z(q
uaat8 sr srualsz(s rredar uerlerurupru aql
Io
JJuel
-rodrur
pue
J)uJtsrxa eql
Jo
uollp)pur
uV
'saseq
0I>
1e
'q8noql 'uoqs
.{1a,rqe1ar
sde.up are saqrled aq1
'a8eruep
ratye
uorsal
q)eJ
le
pazlsaqtur{sal
vN(
Jo
lunolue
aql
ur
LlrauaSoJeleq
Moqs sller uerlpruruew
'(rredag
pue
uortdursup{ uaeMlJq uortf,Juuo)
y
'ZI'VZ
uorlJJS aas) aseraru,{1od
y51g qtrzn
pelerJosse
roDeJ
uorldrJJSupJl e
;o
luauodruor
e sr
'dats
uorsrf,ur aq1 roJ
pa.rrnba:
JseJrleq
p
sI
qJIqM
'uralo.rd
€peg
rql'aseraruLlod
VNU
pup
snleredde
rredar aql ueaMlaq uorl)auuoJ
f,Il
-srueqJau
e eq ol sreadde Jsnel JqJ
'(uortdrns
-upJt
ot
luarurpadrul
aql Sur,rourar)
pa.rredar
,{llBrtuaraJard
sr
puerls pJqrr)supJt
eqr
lpql
sr aruanbasuoJ JqJ'parredar.dlertuara;ard
are
saua8
anrpe
Llpuo4dursue4l'uorldtnsueJt
qlIM
uorDauuoJ slr sr
lseaL
ur
pezrJeppreq)
lsaq
uJJq
seq
tpqt
rredar
Jo
eJnleal Surtsaratur
uV
'sIorrJ
aJnp
-oJlur
01 auord aJour JJp z(aql tanarrzroq
'a8e
-uep
Jo
salrs Jaqto
q8norqr
alerqdar.{aqr
uaq6
'Jarurp
1-1
e alrsoddo rred
v-v
ue ef,npoJlul
,(laternrre Laql :srarurp
auru,{qt
te
auord
-JorJa
lou
JJp rJllel aqr
leql
sr saru.dzua
lrlo
-Lrelna
pup
IerJJlJ€q
aql ueeMtaq
JJUJJJJJIp
y
'saseraruz(lod
VN(
uorsJlsupJl
pJIIpJ
serull
-eruos
are.r{.aq1
'3o1oruoq
ueunq Jql JoJ
sapo)
l€ql
AdX
aua8 aqt
pue
'aa$naln'S
Jo
Lr
JSPJeru
-,{1od
ypq
roy Surpor aua?
ggqyy
erqt'qn
'E
ur
A
pue
AI
saserau.dlod
vNC
roJ apor
leql
saua8
e)nwn
pue
gury
aqt
dq
pelJuuepl
sr salrs
pa8e
-rupp
lp
lerJalpru
areldar 01
vNe
Surnsaqtuls
ut
pJ^lo^ur
saseraru,(1od
vNo
Jo
.dpueyradns
y
'suorlJeJJ
SututoI
ygq
snoSoloruoquou ur
luerJrJep
sl
tnq
'uolleu
-rqruo)er
snoSolouroq
q
tuer)rJJp lou
sr dnor8
-qns
srql
isluelnu
esax
pue
'I
Isaw
'6s1va
rtI
pasPeJJur
aJe sJler uorleurquorar
'lserluor
dg
'tuauelrJ
eqt ot
Lileuuanbas
purq
ueql
TSpeg
pup
'SSppU 'ZEpeu
'luJrrrplrJ
uralordoalrnu
e ruroJ 01
yNCI
papuerls-a18urs
rql ot Surputq
.dq ssarord
aqt satprtrur
IspeU
'slrel
pepuprls
-a13urs
alBraua8 01 spue aJrJ eql uo
peDe
seq
asPalJnuoxe ue Jeuy
'>leeJq
puPJls-elqnop e
]e
xaldruor uralordrllmu
e uroJ suratord
peg asaql
'
LSOW
pue'SSOW',TSOW',
I SOW')SOW
uI
uorteurquo)JJ
)rlolrlu
uI uoltJnper
e Lq uaas
se
'uorteurqruoJal
snoSoloruoq
sDeJJe
dnorE
-qns
JUO
'saddlouaqd
tuplnru
uI JJUeJaJJIp
p
z(q sdnor8qns oMt
otul
papplp
st dnor8
ZEq1v6
eq1'(srusrueq)alu
J>lrl-uolleulqruoJJJ
qlIM
pauraruor)
dnofi
TEgyy
Jql
pue
'(rredar
uolteJ
-qdarrsodro;
parrnbar) dnofi
geW
aqt
'(rredar
uorsrJxJ
ur
pa,r.lonut)
dnor8
tery
eql
dq
pag
-lluepr
'aegnaln's
tseau(
aql u1 sauaE
rrcdar
1o
sdnor8
lerauaE
JaJql aJp erJqJ
'uollpper
ol
,{11
-AltrsuJs
Jreql
Jo
JnIJIA
,{q
tseaz(
ut ,{.lerqaua8
pJZrJal)erPqJ ueeq a,req
Laqt
jsuoll)unJ
rtedar
ur
penlolur
are
lpql
saua8
arB saua8
gyy
'sJseasrp
ueunq
qtIM
pe^lo^ur
eJe sruals.{s
esaql sesef,
IPrJAas
ur
pup
's11ar
ruoLre>1na
raq8tq
ut suBdralunor
lJJrrp
JApq
lsea,{.
ur
punoJ
srualsz(s
rredar aql
;o
.{uew
'uoueurqrxoJal
snoEoloruoq
s1 ulatord
ra;
-suert-pueJls
aqt
Jo
uoll)unJ
ulPru Jqt
'1seaf,
u1
'v)eg
ol
tredralunor
eq1
sI
I Epp5
araq^tr
'1sea,,{.
ur aJe
srualsr(s rrlor{re>1na
pazIJaDPJPqJ
lseq
JqJ'srusrupEro
yo
a8uer
JpIM
p
ol
uoruruoJ eJP
rlor'EUrpezruSorar
suourunJ
JIEdar;o sa&h
aq1
'parredar
r\11e4uara;ard are saua6 anqre
r{l1euot1dulsuet1
o
'
us!ueqleu
I
Jeoal-u0JsJlxa
ueunq e
sapurord
HIIII
lo]teJ uoqdulsuell
eql
pue
slrnpord
4y
6urpnpur
sutalold;o
xalduor
y
r
'sauaD
rredar
1e.re,ras;o
euo fue ut
suoqelnui
fq pasner
aspesrp
upurnq
p
s1
(4X)
urnsoluauEtd
euraporaX
o
'sualsr\s
rteda.t
roJ opol
1eq1
sauab ut are
'sadflouaqd
ent11suas
uorlelppr
[q
pagquepL
'suoqP]nul
6yy
pear\
aq1
o
a^PH
qlal
lqonrPlnl
'sz(emqted
rreda: eqt ur
vJeU
Jo
sDaIJa
DaJIpuI
pue
Derrp
aql 3u113ue1uesrp
uI
InJesn
Jq
plno&r
'asuodsar
SOS
aq] J)npul
or
^lITIqp Jqt
Dplul
3unea1
lnq
uorlJunJ
uolleulquoleJ
aql 3utlen
-llJpur
'uollelnlll
Jo
adz(l
JSJJAJT
Jt{J
'VNO
alp
-ueq
ot
vrrg
Io
Irlgqe aqt sDeJJe
uopetnru
Jo
ad,{1 raqtra1q
'pJlelrDe
aq ot.hrlqe
JI{t
qsuoqe
suorlPlnru
raqlo
'alels
pJlP^IlJe aql ut r{lsnoau
-eluods
surpurar
VJau
esneJeq
r{lqeqord
'tuaru
-1ear1
Suonput
lnoqll,lr
Jn)Jo 01
asuodsar
ggg
Jql s1\^ollp
uollplnlu
Iwv)ar
aq;'luapuadaput
,{1a,rrte1a:
JJe
v)JU
Jo
sJIlInIl)P
o.ttl
eqJ
'(VreU
Jo
uoltelltJe)
roter
-rpur
arues
aq1 o1 Surpuodsar
Iq ruatsds
re1n11ar
aql
oluo Surpeq.{83rd
st uoqrnput
a8eqdord
sualsns
rrPdou
pa^Jasuol

.UOIlEIPPJ
O] JAIIISUJS
JJOIU
SIIJJ ]IlOAIE>INE
IJP
-uar
^etu
sluJuodruot
JsJql
Jo
LuB
ur suortelnw
't)lUX
ulatord Jqt
qlrm
uortlunluol
ur suoll
-JunJ
q)rqM
n1
aseS[
VNO
Jq1
,,{q
ualelrapun
sr
spuJ
pepuprts-JIqnop
aqt
yo
SururoI
lenpe
aq1
'uMou>l
lou
sr suorsnrlord papuerls-a13urs
3ur
-ureruar
Lue
ur sllrJ
ter{l
,(t!rpe
aseraul,lod
y\Iq
aq1
'saua3
urlnqolSounurur
Jo
uorteurqurof,Jr
.dq
paterauaS
surdrreq
Jqt aneall
pup
spue
Sufueqrarro
urJl
qloq
ue)
pue
'sJrlrnrlf,e
espJIJ
-nuopuJ
pue
eseJIJnuoxJ qloq
seq uJoJ
palp^
-rpe
s1r ut
q)rq,t,r
'srruJuv
ulJtord
aql sr sta8rel
JsJr{t
Jo
aug
's1a8re1
uralord
alelLroqdsoqd
ot
vN(
^q
prlenurp
sr
qrrqM
'(sr)a-VNq)
aseurl
uralo:d
luapuadap-vNO
Jq1 sr
tuauodruor
,{."I
V
'ruJqt
uo
]Je
ot sarur(zua
Jeqto sn,rollp
pue
raqlaSol
spuJ
aqt sploq
tpqt
ploJJp)s
e uroJ
z(aq1
'ggn;
pue
O/n>I
suralord
Jql
Jo
Surlsrs
-uof,
reurrpoJJlaq
e,{q spua
ua>loJq Jql
Jo
uorlru
-8orar
sr a8els
lsJrJ
JqI'uorteurquroJJJ
eunrurur
pue
1EHN
qtoq
ur ssaro.rd
aqt
sJ>leuJpun xald
-ruor
aru,{zue
arues
aqJ
'i.ri:",.}.-
}s-*E:i
ur
pJZrJ
-ptuLuns
JJp
l-lHN
ur
pJ^lo^ut
sd:ts aq1
'raq1a3o1
spue
tunlq
ar{t
Surte8
-II
Jo
slslsuor
pue'(1911p)
8u1uro[-pua
snot
-oloruoquou
pJIIEJ
sr
s>leJrq asaql
ledar o1
rusrupr{Jaru ro[Bru
aqI'uor]pperrr
Lq'aldruexe
JoJ-VN11
o1 a8euep
Jo UnsJr
Jqt
sp JntJo
osle Laql
'(uorunag
pue
a8elearg azdlete3
surJlord
gy6
rql
'0I'€Z
uort)es aas)
saua8 uqn
-qolSounrurur
Jo
uorleurquroJJr
Jql
ur elerpJru
-JJluI
up JJe
pup
uorleurquorar
snoSolouroq
;o
ssarord aql
alerllur Laql
'saruelsrunJJrJ
snorJPA
ur sllal ur
rnf,)o s>lpJJq puprls-Jlqnoq
'saseasrp
upunq asnpl
r\emqled
EHN
aql ur suorlplnlrl
o
.VNO
xoldnp
J0
spuo
lunlq
alebrt
uer
r\emqled gHN
agl
o
s)PerB
puPrls-alqno0
srredau
sualsf5 rredaX
97
UlldVHl
gl9
paBeruep
aql SurpnlJur
qJleJls
papuerts-a13urs
aq1
'saua8
dX
trq
pJpoJ
srserlJnuopua dq
uorsel eqt
Jo
Jprs raqtrJ
uo Jppru JJp sJBpApelJ
uaqJ
'uortdrJf,suPrl
Surrnp aseraru,{.10d
yNg
Lq
paralunoJur
sr
teqt
vNc
pa8eruep
;o
rreda:
aqt
qlIM pJ^lolur
sr
qJrqM pup
'saua8
4y
prarras
;o
snnpord Jqt sJpnlJur
qrlqm
'xalduror
a8rel
e
Jleslr
'HII{J
roDpJ
uoltdlnsuerl eql
Jo
.,ilr,rrlre
JSeJrlJq
aqt
riq
ue>lPuepun sr uortJe srql
'alrs
pa8euep
eqt
punorp
dq
OZ-
roJ
punoMun
uer{l
JJe
VNC
Io
spuerls aqJ
'sluJuoduror
str
Jo IeJe
-^JS
Jo
uorJp elrleradoor aql
3ut.1o,rur ursrup
-qJeu
e z(q sdeqrad
'a8euep
Jo
etrs
p
te
vNe
01
spurq xaldruoJ
eqJ
',{e.r,rqled
rredar Jql ut eloJ
sll
sMoqs
tr'*i: =-El1*3i
'sJJrurp
aururtrql
Jo
uors
-rJXe
JoJ alqrsuodsar
sr saua8
gy
Jql
Jo IeJa^as Jo
spnpord
sapnpur
leql
xaldruor
ulatord
y
'sa1o,{re1na
ur
pJsn
,{lapulr
st ,{e,,r,rqred
srql
tpql
Sur,r,roqs
'lsea.{
;o
saua8
OW
eql ur sSolouroq
aneq
Laql
'y-dx
otV-dX
'qrlalls
pasoxo
aq] ereldar o1
pazrseqluAs
sL
vN0
Mou
pue
'uorsal
aqlJ0 aprs raqlra uo
lnl
sosPollnuopue
'a1s
pabeurep
p
te
VN0
spurMun oselrtaq
V
€I"*f
$*{tStj
uats^s
uoruruol
v
.JSE.IEUI
-,{1od
y51q
Jql
Jo
ssol ot
anp Llqerunsard
are
sluelnru/dX
ur JnJJo
teql
sJeJuel ul{S
ZdX
^q
papoJ
sr srqJ
'Jerurp
eqt
lsed
pJJJord
o1 rapro
ur
.dtr.Lrtre
Lr
aserarudlod
vNq
aqt sarrnbar
uorleJ
-[dar
'panorueJ
uJaq
tou
spq
lpql
JJturp
autu.r{qt
e srJlunoJua
uortelrldar
alJqM
sJSpJ
ul
'IUJlueJPIOJJ
P
Jo
srseqluz(s
,{q
pareldal
Jq
uJql ueJ
sJseq
uorsol
aq]
lo
eprs
,9
orll
so^ealc
tccHf/ldx
xrlor{
olqnop eq1 suedo
;.1tt31
saseelcnuopua
sasecrloLl
pebeuep
sr
y11g

Some
of the
genes
for these
proteins
are mutated
in
patients
who
have
diseases due to deficien-
cies in DNA repair.
The
I(u heterodimer is
the sensor that
detects
DNA
damage by binding to the broken
ends. The crystal structure in
Ft{liltL
}i.:.i5 shows
why
it
binds
only to ends. The
bulk of
the
pro-
tein extends
for
about two turns along one face
of DNA
(lower).
but
a narrow
bridge between
the subunits,
located in the center of the struc-
ture, completely encircles
DNA. This means
that
the
heterodimer needs to slip onto a free end.
I(u can bring broken ends
together by bind-
ing
two
DNA molecules. The ability of
I(u het-
erodimers to associate with one
another suggests
that the reaction might take
place
as
illustrated
in iiiirlhli=
.".ii.i*.
This would
predict
that the
li-
gase
would act by
binding in the region between
the bridges
on the individual heterodimers.
Pre-
sumably I(u
must
change
its
structure
in order
to be
released from DNA.
Deficiency
in DNA repair
causes
several
human diseases.
The
common
feature is that
an
inability to repair double-strand breaks
in
DNA leads
to
chromosomal
instability. The insta-
bility
is revealed by chromosomal aberrations,
which are associated with an
increased rate of
mutation, which in turn leads to an
increased
susceptibility
to cancer in
patients
with the dis-
ease. The basic cause can be
mutation in
path-
ways
that control DNA repair or
in
the
genes
that code
for
enzymes of the
repair complexes.
The
phenotypes
can be very similar, as
in the
case of Ataxia telangiectasia
(AT),
which
is
caused
by failure of a cell cycle checkpoint
path-
way, and Nijmegan
breakage syndrome
(NBS),
which
is
caused
by a mutation of a repair
enzyme. One
of the lessons that we
learned
from characterizing
the repair
pathways
is that
they
are conserved
in mammals,
yeast,
and
bacteria.
The
recessive human disorder of Bloom's
syndrome
is caused by mutations
in a helicase
gene
(called
BLM)
that
is homologous to recQ
oI E coli.
The mutation results in an
increased
frequency of
chromosomal breaks and sister
chromatid exchanges.
BLM associates with other
repair
proteins
as
part
of
a large complex. One
of the
proteins
with which
it interacts is hMLHl,
a
mismatch-repair
protein
that is the human
homolog of bacterial mutL.The
yeast
homologs
of these
two
proteins,
Sgsl and
MLHI, also asso-
ciate,
identifying these
genes
as
parts
of
a well-
conserved
repair
pathway.
Nijmegan breakage
syndrome
results
from mutations
in a
gene
coding for a
protein
i:*li*:i:
.r:i-:"ij'+ Nonhomologous
end
joining
requires recog-
nition
of the
broken ends,
trimming
of overhanging
ends
and/or
filting, fotlowed
by tigation.
i:t{iLli.i{: J{.i,.ilii
The Ku70-Ku80
heterodimer
binds along
two turns of the
DNA doubte
hetix and
surrounds
the
helix
atthe center ofthe
binding
site.
Photos
courtesy
ofJonathan
Go[dberg,
MemoriaI
Sloan-
Ketteri
ng Cancer
Center.
20.1.2
A Common
System
Repairs
Doubte-Strand
Breaks
517

F:*il€i
!ri.l5
If two heterodimers
of
Ku
bind to DNA, the
distance
between
the two
bridqes
that encircte DNA is
-12
bp.
(variously
called
Nibrin
,
p95,
or NBS 1)
that is
a component
of the Mre I
I /Rad50 repair
com-
plex.
Its involvement
in repairing
double-
strand
breaks is
shown by
the formation
of
foci
containing
the
group
of
proteins
when human
cells
are irradiated
with
agents
that induce
double-strand
breaks. After
irradiation,
the
kinase
ATMP
(coded
by the ,4T
gene) phos-
phorylates
NBSl; this
activates
the complex,
which
localizes
at
sites of DNA
damage.
Sub-
sequent
steps involve
triggering
a
checkpoint
(a
mechanism
that
prevents
the
cell cycle from
proceeding
until
the damage
is repaired)
and
recruiting
other
proteins
that
are required
to
repair
the
damage
@
Summary
Bacteria
contain
systems
that maintain
the
integrity
of their
DNA
sequences in
the face
of
damage
or errors
of replication
and that distin-
guish
the DNA
from
sequences
of a foreign
source.
Repair
systems
can recognize
mispaired,
altered,
or
missing
bases in DNA,
as well
as other
structural
distortions
of
the double
helix. Exci-
sion
repair
systems
cleave
DNA
near a site
of
damage,
remove
one strand,
and
synthesize
a
new
sequence
to replace
the excised
material.
The
Uvr system
provides
the main
excision-
repair pathway
in
E. coli.
The
dam system
is
involved
in
correcting
mismatches generated
by incorporation
of incorrect
bases
during repli-
cation
and
functions
by
preferentially
remov-
ing
the
base
on the
strand
of DNA
that is not
methylated
atthe
dam target
sequence.
Eukary-
otic homologs
of the E.
coli
MrttSL
system are
involved
in
repairing
mismatches
that
result
from
replication
slippage;
mutations
in this
path-
way
are
common
in
certain types
of
cancer.
CHAPTER
20 Repair
Systems
Recombination-repair
systems
retrieve
information
from
a DNA
duplex and
use it to
repair
a sequence that has
been
damaged
on
both strands. The
RecBC
and RecF
pathways
both act
prior
to RecA,
whose
strand-transfer
function is involved
in all
bacterial recombina-
tion. A major
use of recombination-repair
may
be to recover
from the
situation
created
when
a replication
fork
stalls.
The
other capacity
of recA is
the ability
to
induce
the SOS response.
RecA
is activated
by
damaged DNA
in an unknown
manner.
It
trig-
gers
cleavage of the LexA
repressor protein,
thus releasing
repression
of many loci,
and
inducing
synthesis of the
enzymes
of both
exci-
sion
repair
and recombination-repair
pathways.
Genes under
LexA control
possess
an
operator
SOS
box. RecA also
directly activates
some repair
activities. Cleavage
of repressors
of lysogenic
phages
may induce
the
phages
to
enter the lytic
cycle.
Repair
systems can
be connected
with tran-
scription
in both
prokaryotes
and
eukaryotes.
Human
diseases
are caused
by mutations
in
genes
coding for
repair
activities
that are
asso-
ciated
with the transcription
factor
TFIIH.
They
have
homologs in
the RAD
genes
of
yeast,
which
suggests
that this repair
system
is widespread.
Nonhomologous
end-joining
(NHEJ)
is a
general
reaction
for repairing
broken
ends in
(eukaryotic)
DNA.
The I(u heterodimer
brings
the
broken ends
together so
they can
be ligated.
Several
human
diseases
are caused
by muta-
tions in
enzymes
of this
pathway.
References
Repair
Systems
Correct Damage
to
DNA
Reviews
Sancar, A.,
Lindsey-B
oltz, L. A.,
Unsal-Kagmaz,
K.,
and Linn,
S.
(2004).
Molecular
mechanisms
of
mammalian
DNA repair
and
the DNA
damage
checkpoints.
Annu.
Rev. Biochem
73,
)9-85.
Wood, R. D.,
Mitchell, M.,
Sgouros,
J.,
and Lin-
dahl, T.
(2001).
Human
DNA
repair
gene
s.
Scl-
ence 29I, 1284-1289.
Excision-Repair
Pathways
in Mammalian
Ce[ts
Reviews
Barnes,
D. E. and
Lindahl,
T.
(2004t.
Repair
and
genetic
consequences
of endogenous
DNA
base damage
in mammalian
cells. Annu
Rev.
Genet )8,445-476.
518

McCullough, A. I(., Dodson, M. L.,
and
Lloyd, R.
S.
(1999\.
Initiation
of base excision repair:
gly-
cosylase mechanisms and structures. Annu
Rev. Biochem
68,
255-285.
Sancar,
A., Lindsey-BoItz,L. A.,
Unsal-I(agmaz,
I(.,
and Linn, S.
(2004).
Molecular mechanisms of
mammalian DNA repair and
the
DNA damage
checkpoints.
Annu Rey. Biochem. T),
)9-85.
Resea
rc h
ICungland, A. and Lindahl, T.
(1997lr.
Second
pathway
for
completion oI human DNA base
excision-repair:
reconstitution
with
puri{ied
proteins
and requirement for DNase IV
(FENI).
EMBO J. 16,
j)4r-3348.
Matsumoto,
Y.
and
I(im, K.
(1995).
Excision
of
deoxyribose
phosphate
residues by DNA
poly-
merase beta during DNA repair.
Science
269,
699-702.
Base Ftipping Is Used by Methytases
and Glycosytases
Resea rch
Aas, P. A., Otterlei,
M., Falnes, P A.,
Vagbe, C.
B.,
Skorpen,
F., Akbari, M.,
Sundheim,
O., Bjoras,
M., Slupphaug, G., Seeberg, E., and
I(rokan,
H. E.,(2003). Human and bacterial oxidative
demethylases
repair alkylation damage in
both RNA and
DNA.
Nature
421,859-863.
Falnes, P. A., Johansen,
R. F.,
and Seeberg,
E.
(2002\.
AlkB-mediated oxidative demethyla-
tion
reverses DNA damaee in E. coli. Nature
4r9, t78-182.
ICimasauskas, S.,
Kumar,
S.,
Roberts, R.
J.
and
Cheng,
X.
(1994\.
HhaI methyltransferase
flips
its
target
base out of the DNA helix. Cel/
76,357-J69.
Lau, A.
Y.,
Glassner,
B.
J.,
Samson, L. D., and
Ellenberger,
T.
(2000).
Molecular basis
for
dis-
criminating between
normal
and
damaged
bases
by the human alkyladenine
glyco-
syiase,
AAG. Proc Natl Acad. Sci. USA 97
,
tj57)-t)578.
Lau,
A. Y., Scherer, O. D., Samson, L., Verdine,
G.
L., and Ellenberger,
T.
(1998).
Crystal
struc-
ture of a
human alkylbase-DNA repair
enzyme complexed
to DNA: mechanisms
for
nucleotide
flipping and base excision. Cell
95,
249-258.
Mol, D. D. et al.
(1995).
Crystal structure
and
mutational analysis of
human uracil-DNA
gly-
cosylase:
structural basis
for
specificity
and
catalysis. Cell
80, 869-878.
Park, H. W.,
ICm,
S.
T.,
Sancar,
A., and Deisen-
hofer,
J.
(1995).
Crystal structure
of DNA
photolyase
from E. coli. Science
268,
t866-1872.
Sawa,
R. et al.
(
I 99 5
)
. The structural basis of spe-
cific
base-excision
repair by uracil-DNA
glyco-
sy\ase.
Nature 373,
487-49).
Trewick,
S. C.,
Henshaw,
T. F.,
Hausinger,
R. P.,
Lindahl, T., and
Sedgwick,
B.
(2002).
Oxida-
tive demethylation
by
E coli AlkB
directly
reverts DNA base damage.
Nature
419,
t74-r78.
Vassylyev, D. G. et al.
(1995). Atomic
model of a
pyrimidine
dimer
excision
repair
enzyme
complexed with
a DNA substrate:
structural
basis for damaged
DNA
recognition.
Cell 8),
773-782.
Error-Prone
Repair
and
Mutator
PhenotYPes
Resea
rc h
Friedberg, E.
C.,
Feaver,
W. J.,
and Gerlach,
V.
L.
(2000).
The
many
faces of
DNA
polymerases:
strategies for
mutagenesis
and
for mutational
avoidance. Proc
Natl.
Acad. Sci. USA
97,
5681-568).
Goldsmith,
M., Sarov-Blat,
L., and
Livneh,
Z.
(2000).
Plasmid-encoded
MucB
protein is a
DNA
polymerase
(pol
RI)
specialized
for
lesion
bypass in the
presence oI
MucA,
RecA, and
SSB.
Proc. Natl
Acad Sci.
USA
97,
11227-tt23t.
Maor-Shoshani,
A., Reuven,
N.
B., Tomer,
G., and
Livneh, Z.
(2000\
. Highly
mutagenic
replica-
tion by
DNA
polymerase
V
(UmuC)
provides
a
mechanistic basis
for SOS
untargeted
muta-
genesis.
Proc. Natl.
Acad
Sci. USA97,
565-570'
Wagner, J.,
Grtrz,
P., Kim, S.
R.,
Yamada,
M., Mat-
sui,
K., Fuchs,
R. P., and
Nohmi,
T.
(1999).
The dinB
gene
encodes
a
novel
E. coli
DNA
polymerase, DNA
pol
IV
involved
in mutagen-
esis.
Mol. Cell
4,281-286.
Controlting
the
Direction
of
Mismatch
RePair
Review
I(unkel,
T. A., and
Erie, D.
A.
(2005). DNA mis-
match repair.
Annu
Rev.
Biochem.74,681-710'
Resea rc h
Strand,
M., Prolla,
T. A., Liskay,
R. M., and
Petes,
T. D.
(t993).
Destabilization
of
tracts of
simple
repetitive
DNA
in
yeast
by
mutations
affecting
DNA mismatch
repair.
Nature )65,274-276.
Recombination-Repair
Systems
in E' coli
Review
West,
S. C
(1997i. Processingof
recombination
intermediates
by the
RuvAB C
proteins.
Annu.
Rev. Genet. )
l, 2lJ-244.
Resea
rch
Bork, J. M. and
Inman,
R.
B.
(2001). The RecOR
proteins
modulate
RecA
protein function
at 5'
ends
of single-stranded
DNA.
EMBO J.20,
7)t)-7)22.
References
519

@
Recombination
Is an Important
Mechanism
to Recover
from
Reptication
Errors
Reviews
Cox, M.
M.,
Goodman, M. F., Kreuzer.
I(. N.. Sher-
ratt,
D.
J., Sandler, S.
J., and Marians, I(.
J.
(2000).
The importance
of
repairing
stalled
replication
forks.
Nature
404, )7-41 .
McGlynn,
P
and Lloyd, R.
c.
(2002).
Recombina-
tional repair
and restart
of damaged
replica-
tion forks.
Nat Rev.
Mol.
Cell
Biol.
), 859-870.
Michel,
B,, Viguera,
E., Grompone,
G.,
Seigneur,
M.,
and Bidnenko,
V
(2001).
Rescue
of arrested
replication
forks
by homologous
recombina-
tion. Proc
Natl Acad
Sci. USA 98,
8l8l-8188.
Resea rc
h
Courcelle,
J. and Hanawalr,
p
C.
(2003).
RecA-
dependent
recovery
of arrested
DNA replica-
tion forks.
Annu. Rev.
Genet 37,
6ll-646.
I(uzminov,
A.
(2001
).
Single-strand
interruptions
in
replicating
chromosomes
cause double-
strand
breaks . Proc.
Natl. Acad.
Sci. USA 98.
8241-8246.
Rangarajan,
S., Woodgate, R.,
and
Goodman, M. F.
(19991.
A
phenotype
for enigmaric
DNA
poly-
merase II:
a
pivotal
role for
pol
II in replication
restart
in
UV-irradiated
Escherichia
coli Proc.
Natl Acad
Sci. USA 96, 9224-9229.
RecA Triggers
the
S0S System
Research
Tang,
M. et
al.
(
I 999).
UmuD'2C is
an error-prone
DNA polymerase,
E.
coli
polY.
Proc. Natl. Acad.
Sci. USA 96,
8919-8924.
W
Eukaryotic
Celts Have
Conserved
Repair
Systems
Reviews
Iftogh,
B.
O. and Symington,
L.
S.
(2004).
Recom-
bination
proteins
in yeast.
Annu Rev.
Genet.
)8,2)j-27t.
Prakash,
S. and Prakash,L. (2002\.
Tfanslesion
DNA
synthesis
in eukaryotes:
a one- or two-
polymerase
aff.air.
Genes Dev. 16,
1872-188).
Sancar,
A.,
Lindsey-B
oltz, L.
A., Unsal-I(agmaz,
K.,
and Linn,
S.
(2004).
Molecular
mechanisms
of
mammalian
DNA repair
and
the DNA damage
checkpoints.
Annu
Rev. Biochem
7),
j9-85.
Resea
rc h
Friedberg,
E.
C.,
Feaver,
W. J., and
Gerlach, V.
L.
(2000).
The
many
faces
of DNA
polymerases:
strategies
for mutagenesis
and for mutational
avoidance.
Proc
Natl. Acad.
Sci.
USA 97.
568 l-t683.
Johnson,
R. E
,
Prakash,
S., and
Prakash,
L.
(1999).
Efficient
bypass
of a
thymine-thymine
dimer
by
yeast
DNA
polymerase,
Pol eta.
Scl-
ence 28),
1001-1004,
CHAPTER
20 Repair
Systems
Rattray, A.
J. and Strathern,
J. N.
(2003).
Error-
prone
DNA
polymerases:
when making
a mis-
take is
the only way to
get
ahead. Annu.
Rev.
Genet. 37, )l-66.
Reardon,
J. T. and Sancar,
A.
(2003).
Recognition
and repair
of the cyclobutane
thymine
dimer,
a major
cause oI skin cancers,
by the
human
excision nuclease.
Genes Dev.
17
,
2*9-2551.
Wolner, 8.,
van I(omen,
S., Sung, P.,
and Peterson,
C.
L.
(2003).
Recruitment
of
the recombina-
tional repair machinery
to a DNA
double-
strand break in
yeast.
Mol
Cell 12,221-232.
A
Common System
Repairs
Doubte-Strand
Breaks
Review
D'Amours,
D. and
Jackson, S. P.
(2002).
The
Mrel I complex:
at the crossroads
of DNA
repair and
checkpoint signalling.
Nat. Rev. Mol.
Cell Biol
),317-327.
Resea rc h
Carney,
J.
P.,
Maser, R.
S., Olivares,
H., Davis,
E. M.,
Le Beau, M.,
Yates,
J.
R.,
Hays, L.,
Morgan,
W. F., and Petrini,
J. H.
(1998).
The hMrel
I /hRad5O
protein
complex
and
Nijmegen breakage
syndrome:
linkage
of
double-strand
break repair
to the cellular
DNA
damage response.
Cell 93, 477
486.
Cary, R. B., Peterson,
S. R.,
Wang,
J., Bear, D.
G.,
Bradbury,
E. M., and
Chen, D.
J.
(1997).
DNA
looping
by Ku and
the DNA-dependenr pro-
tein kinase.
Proc. Natl
Acad.
Sci. USA 94.
4267-4272.
Ellis,
N. A.,
Groden, J., Ye,
T. 2.,
Straughen,
J.,
Lennon,
D. J.,
Ciocci, S., Proytcheva,
M., and
German,
J.
(1995).
The Bloom's
syndrome
gene
product
is homologous
to RecQ
heli-
cases.
Cell 83, 655-666.
Ma, Y., Pannicke,
U., Schwarz, I(.,
and Lieber,
M.
R.
(2002).
Hairpin
opening
and
overhang
pro-
cessing
by an Artemis/DNA-Dependent
pro-
tein kinase
complex in
nonhomologous
end
joining
and
V(D)J
Recombinarion.
Cell 108,
78t-794.
Ramsden,
D. A.
and
Gellert, M.
(199S).
I(u
prorein
stimulates
DNA
end
joining
by mammalian
DNA
ligases:
a direct role
for I(u
in repair
of
DNA
double-strand
breaks. EMBO
J.
17,
609-6t4.
Varon,
R. et
al.
(1998).
Nibrin, a novel
DNA
dou-
ble-strand
break repair protein,
is
mutated
in
Nijmegen
breakage
syndrome.
Cell 93,
467-476.
Walker,
J. R.,
Corpina, R. A.,
and
Goldberg,
J.
(2001
)
.
Structure of
the I(u heterodimer
bound
to DNA
and its implications
for
double-
strand break
repair.
Nature 412,
607-614.
520