Куценко С.А. Основы токсикологии
Подождите немного. Документ загружается.

Общая характеристика реакций конъюгации ксенобиотиков представлена в таблице 4.
Таблица 4. Характеристика основных реакций конъюгации ксенобиотиков
Реакция Присоединяемый агент
Функциональная группа
ксенобиотика
А. Реакции, протекающие при участии активированных форм присоединяемых агентов
Конъюгация с
глюкуроновой кислотой
УДФ-глюкуроновая
кислота
-ОН; -СООН; NH
2
;
-NR
2
; -SH; -CH
Конъюгация с глюкозойУДФ-глюкоза -ОН; -SH; COOH; =NH
Сулфатация ФАФС -ОН; -NH
2
; -SH
Метилирование S-аденозилметионин -ОН; -NH
2
Ацетилирование Ацети КоА -ОН; -NH
2
Детоксикация цианида Сульфон-сульфид -CN
-
Б. Реакции, протекающие при участии активированных форм ксенобиотиков
Конъюгация с
глутатионом
Глутатион
Ареноксиды; эпоксиды;
галогенированные алкильные и
арильные углеводороды
Конъюгация с
аминокислотами
Глицин; глутамин;
орнитин; таурин; цистеин
-СООН
(J. Caldwell, 1979)
В ряде случаев, в ходе метаболизма ксенобиотиков во ll фазе также образуются токсичные
продукты (токсификация).
4.1. Ацетилирование
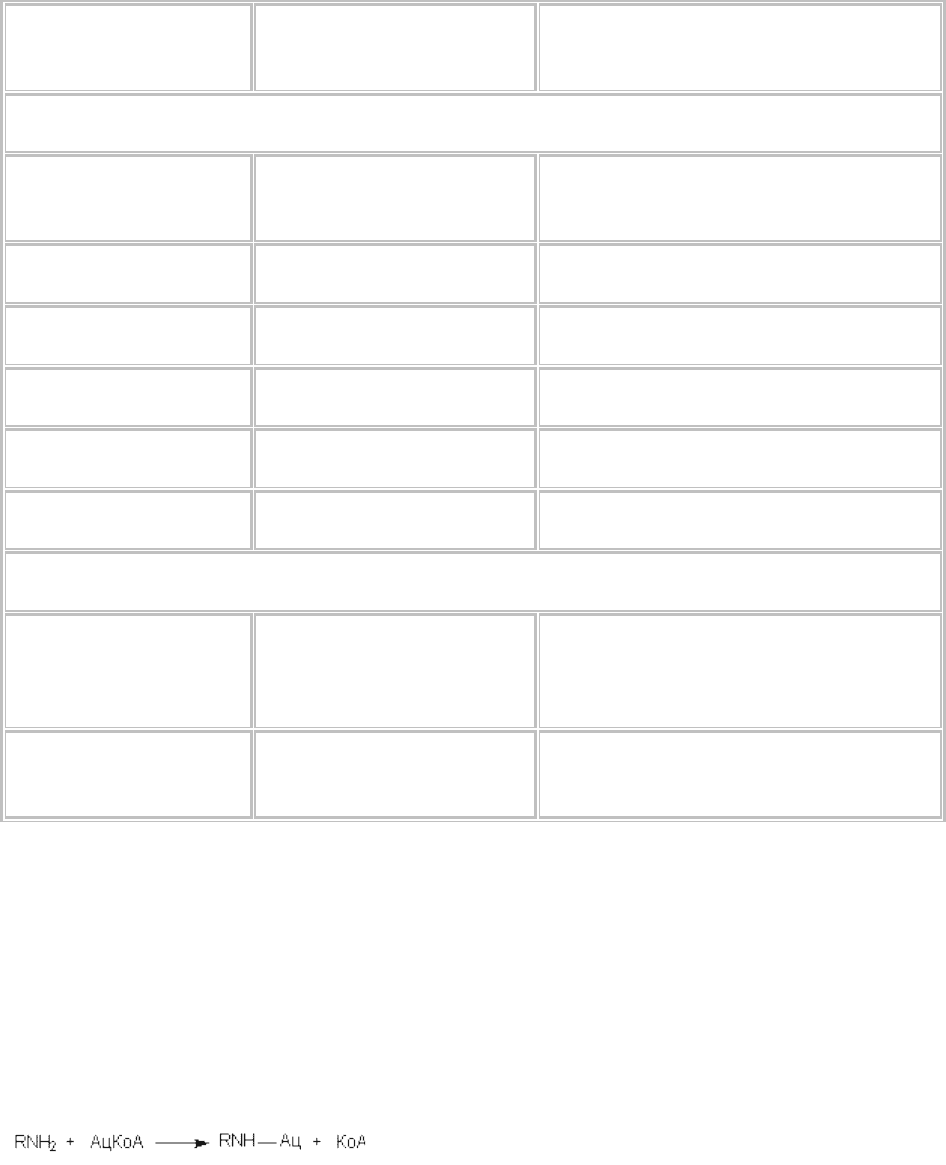
Аминогруппы ароматических соединений часто подвергаются ацетилированию. Уксусная кислота
переносится на аминогруппу в форме ацетил-КоА с помощью соответствующих трансфераз, в
частности - ацетил-КоА:амин-N-ацетилтрансферазы.
Ацетилированию могут подвергаться ариламингруппы, сульфамидные группы, алифатические
амины, группы гидразина. У людей выражены генетически обусловленные различия в способности
к реакции N-ацетилирования. Замедленное ацетилирование отмечается у гомозиготных по
рецессивному аллелю индивидов. Высокая активность ацетилирования отмечается у
гетерозиготных индивидов или людей, гомозиготных по доминантному аллелю.
4.2. Другие реакции ацилирования
Не только уксусная кислота, но и другие органические кислоты способны превращаться в
организме в активную форму, вступая во взаимодействие с КоА (жирные кислоты, карболовая
кислота, бензойная кислота, фенилуксусная кислота и др.). В этой форме вещества вступают в
реакцию взаимодействия с соединениями, содержащими аминогруппу (глицином, глутаматом), с
образованием конъюгатов. Так, известно, что при поступлении в организм бензойной кислоты с
мочой выделяется гиппуровая кислота. Гиппуровая кислота представляет собой конъюгат
бензойной кислоты с глицином. В данном случае эндогенная молекула глицина выступает в
качестве акцептора, с которым связывается бензойная кислота, активированная КоА. В организме
человека активированная форма фенилуксусной кислоты связывается с глутаматом.
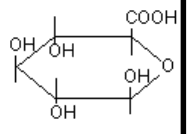
4.3. Конъюгация с глюкуроновой кислотой
Глюкуроновая кислота (рисунок 11) имеет большое значение в механизме биотрансформации
ксенобиотиков.
Рисунок 11. Глюкуроновая кислота
Она активно присоединяется к молекулам алифатических и ароматических спиртов, органических
кислот, серосодаржащих соединений. Процесс конъюгации приводит к образованию эфиров
глюкуроновой кислоты - глюкуронидов.
В реакцию конъюгации глюкуроновая кислота вступает в активной форме
уридиндифосфоглюкуроновой кислоты (УДФГК) и переносится на молекулу-акцептор с помощью
соответствующей трансферазы: УДФ-глюкуронозилтрансферазы (УДФ-ГТ). Энзим
идентифицирован в микросомальной фракции клеток печени, почек, других органов. УДФГК
образуется в процессе взаимодействия глюкозо-1-фосфата с уридинтрифосфорной кислотой
(УТФ) в растворимой фракции цитозоля клеток. УДФ-ГТ индуцируется при поступлении в организм
таких веществ, как фенобарбитал, ПАУ, диоксины, полигалогенированные бифенилы.
Примеры типов реакции глюкуронидирования представлены на рисунке 12.
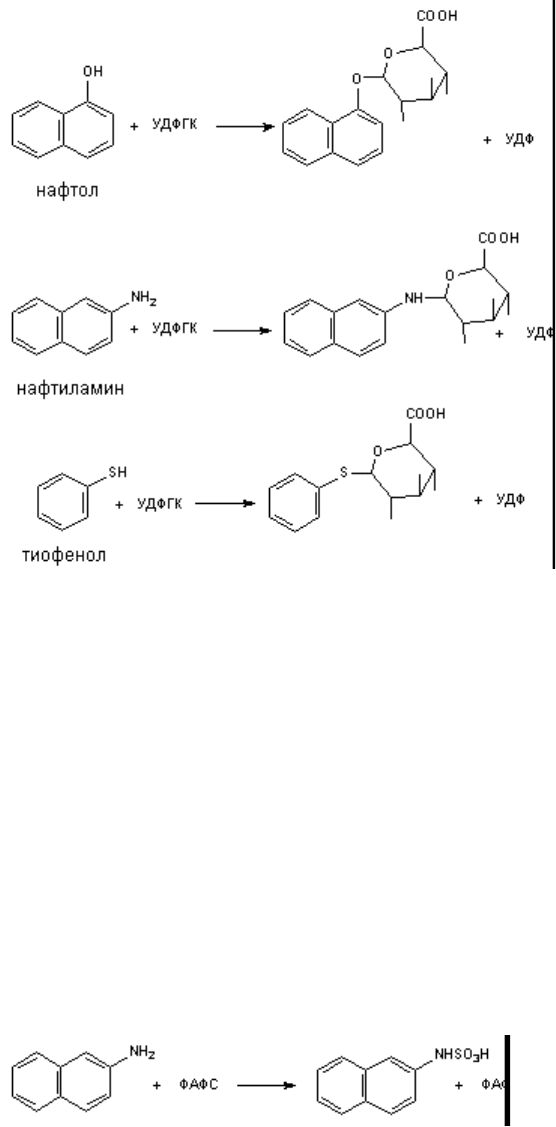
Рисунок 12. Некоторые реакции глюкуронидирования ксенобиотиков
С помощью конъюгации с глюкуроновой кислотой метаболизируют и некоторые эндогенные
вещества, например стероиды и билирубин.
В кишечнике под влиянием глюкуронидазы, энзима кишечной микрофлоры, глюкурониды могут
расщепляться с образованием веществ, способных к реабсорбции и обратному поступлению в
кровь (явление кишечно-печеночной циркуляции ксенобиотика).
4.4. Конъюгация с сульфатом
Различные соединения, содержащие фенольные группы выделяются из организма в виде
конъюгатов с сульфатом. Эндогенные сульфаты могут взаимодействовать также с
ароматическими аминами. Процесс взаимодействия проходит в несколько этапов. На первом
этапе образуется активная форма сульфата - 3-фосфоаденазин-S-фосфосульфат (ФАФС).
Перенос сульфогруппы на молекулу-акцептор (фенол, стероиды и др.) осуществляется с помощью
энзима сульфотрансферазы:
В зависимости от строения молекулы-акцептора в процесс вовлекаются различные
сульфотрансферазы. Энзимы не индуцируются ксенобиотиками. Их активность может быть
угнетена пентахлорфенолом, 2,6-дихлор-4-нитрофенолом. Сульфотрансферазы обладают
относительно высокой субстратной специфичностью. Система конъюгации сульфата локализуется
главным образом в цитозольнй фракции гепатоцитов. Запасы ФАФС в печени незнечительны,
легко истощаются, что при высоких токсических нагрузках приводит к переключению метаболизма
на другие пути, в частности в сторону образования продуктов глюкуронидирования. Сульфатация,
таким образом, является системой с "высоким сродством, но малой мощностью",
глюкуронидирование, напротив - с "малым сродством, но высокой мощностью".
4.5. Конъюгация с глутатионом и цистеином
Органические вещества, содержащие в молекуле лабильные атомы водорода, галогенов и др., в
организме могут взаимодействовать с SH-содержащими эндогенными соединениями: цистеином,
ацетилцистеином, глутатионом и т.д.
Реакции конъюгации восстановленного глутатиона с электрофильными субстратами катализируют
энзимы глутатион-S-трансферазы (GST). Хотя GST обнаружены практически во всех тканях
организма млекопитающих, уровень энзимов в печени - максимален. В этом органе GST
составляет до 10% общего количества цитозольных протеинов. GST - большое семейство
энзимов, в котором набор изоформ часто специфичен для отдельных тканей. Их активность
индуцируется при поступлении в организм различных ксенобиотиков. В подавляющем
большинстве случаев взаимодействие ксенобиотиков с глутатионом приводит к их детоксикации.
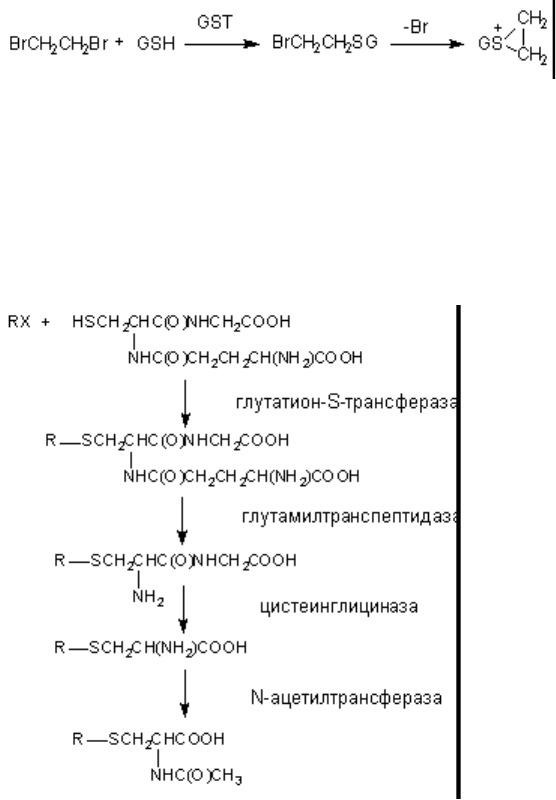
Однако известны примеры биоактивации. Так, конъюгация глутатиона с 1,2-дигалогеналканами
(дихлорэтан, дибромэтан) приводит к образованию продукта: S-(2-галогеналкан), который в
дальнейшем превращается в активный циклический эписульфониум-ион (рисунок 13).
Рисунок 13. Биоактивация дибромэтана при участии глутатион-S-трансферазы
Глутатион, вступивший в реакцию конъюгации с веществом подвергается расщеплению (рисунок
14). В результате из организма выделяется метаболит в связанной с N-ацетилцистеином форме
(меркаптуровые кислоты).
r
Рисунок 14. Взаимодействие ксенобиотика с глутатионом и последующее превращение комплекса
Аналогичным образом происходит взаимодействие ксенобиотиков с цистеином и
ацетилцистеином:

4.6. Метилирование
Для многих веществ, процесс превращения завершается этапом метилирования молекулы.
Метилированию могут подвергаться молекулы, содержащие гидроксильные, сульфгидрильные и
аминогруппы в структуре. В качестве донора метильной группы выступает метионин в форме S-
аденозилметионина (SАМ). Перенос радикала осуществляют соответствующие O-, S-, N-
метилтрансферазы:
Среди эндогенных веществ, таким образом, при участии фермента катехол-О-метилтрансферазы
(КОМТ) метаболизируют адреналин, норадреналин, дофамин. При этом образуются малоактивные
3-метоксипроизводные катехоламинов.
5. Энзимы кишечной флоры
При участии кишечной флоры также возможен метаболизм химических соединений. Действие
бактериальных энзимов сопровождается расщеплением продуктов ll фазы метаболизма,
поступающих в кишечник с желчью, и образованием исходных метаболитов. Этот процесс
проходит при участии гидролаз, разрушающих глюкурониды и сульфаты. Реабсорбция
образовавшихся веществ замыкает цикл внутрипеченочной рециркуляции ксенобиотиков. Кроме
того, анаэробная среда кишечника обеспечивает возможность восстановления некоторых
химических веществ бактериями. Например, восстановление нитроароматических соединений
сопровождается образованием аминов, которые, поступая в печень, вновь подвергается
метаболизму. Бактериальная -глюкуронидаза и нитроредуктаза играют исключительно важную
роль в процессе многоэтапной биоактивации 2,6-динитротолуола (рисунок 15).
Рисунок 15. Взаимодействие печеночных энзимов и энзимов флоры кишечника в процессе
биоактивации канцерогена 2,4-динитротолуола
6. Факторы, влияющие на метаболизм ксенобиотиков
Способность органов и тканей метаболизировать ксенобиотики зависит от набора и активности
энзимов, участвующих в процессе. В значительной степени активность энзимов является
внутренней характеристикой конкретной ткани, определяется генетическими особенностями
организма и зависит от пола и возраста. Дополнительными факторами, порой существенным
образом влияющими на содержание и активность энзимов, являются условия окружающей среды.
Это прежде всего химические вещества, выступающие в качестве индукторов или ингибиторов
энзимов, питание и действие патогенных факторов (таблица 5).
Таблица 5. Факторы, влияющие на метаболизм ксенобиотиков
ЕСТЕСТВЕННЫЕ:
вид, пол, возраст, питание
ИНДУКТОРЫ ФЕРМЕНТОВ:
Барбитураты, полициклические углеводороды, андрогенные стероиды, анаболические
стероиды, глюкокортикоиды и др.
ИНГИБИТОРЫ ФЕРМЕНТОВ:
Метирапон, 7,8-бензофлавон, кобальт, SKF-525 и др.
ПОВРЕЖДЕНИЯ СТРУКТУРЫ ОРГАНА:
Хлорированные углеводороды, тироксин, аллоксан, морфин, гепатотомия,
адреналэктомия, кастрация самцов, голод
6.1. Генетические факторы
Особи одного и того же вида живых существ порой значительно различаются по способности
метаболизировать ксенобиотики. Это во многом детерминировано генетически. Так, в популяции
людей выявляются лица, обладающие пониженной активностью цитохромР-450 зависимых
оксидаз. "Слабые метаболизанты" могут отличаться отсутствием некоторых изоэнзимов,
необходимых для катализа ряда превращений ксенобиотиков.
Полиморфизм метаболизма ксенобиотиков отмечен для процессов ацетилирования
ароматических веществ, содержащих амино-, сульфо-, амидную группу. Лица со слабым
напряжением процессов ацетилирования более подвержены некоторым аллергическим реакциям,
вызываемым химическими веществами. С другой стороны, излишне напряженный процесс
ацетилирования ксенобиотиков может иметь пагубные последствия в результате биоактивации
арилгидразинов.
6.2. Пол и возраст
В опытах на лабораторных животных, в основном грызунах, показано, что половые гормоны
принимают участие в регуляции активности энзимов метаболизма ксенобиотиков и прежде всего
монооксигеназ. Так, взрослые самцы крыс быстрее метаболизируют такие вещества, как
гексабарбитал, аминопирин, аминофенол и т.д. Вместе с тем анилин и его аналоги подвергаются
биопревращению в организме самцов и самок с одинаковой скоростью. Кастрация нередко
сопровождается снижением скорость метаболизма ксенобиотиков. У человека половые различия
выражены не столь существенно.
Отличия метаболизма ксенобиотиков, обусловленные возрастом, наиболее отчетливо
проявляются у новорожденных и лиц пожилого возраста. Хорошо известно, что недостаточно
развитая система метаболизма ксенобиотиков у новорожденных делает их особенно
чувствительными к ряду токсикантов. Токсический процесс может стать следствием как
накапливающихся в организме исходных продуктов, так и промежуточных метаболитов, не
подвергающихся дальнейшей биотрансформации.
В старческом возрасте наблюдается снижение клиаренса ксенобиотиков отчасти обусловленное
понижением интенсивности метаболизма. Нарушение метаболизма ксенобиотиков печенью в
старческом возрасте может являться следствием уменьшения интенсивности печеночного
кровотока, хронических патологических процессов в печени, связанного с возрастом снижения
активности ферментов.
6.3. Влияние химических веществ
Ксенобиотики, поступающие в организм, могут оказывать влияние на процессы метаболизма как
самих этих веществ, так и других соединений, поступающих в организм одновременно или вслед
за ними. Теоретически можно выделить три группы химических соединений, по-разному влияющих
на метаболизм чужеродных веществ:
1. Практически не влияющие на активность энзимов метаболизма;
2. Повышающие активность энзимов - индукторы;
3. Угнетающие активность энзимов - ингибиторы.
Важно иметь в виду, что одно и то же вещество может выступать и как индуктор и как ингибитор
метаболизма другого вещества, в зависимости от того в каком порядке ксенобиотики поступают в
организм - сукцессии или комбинации (см. ниже).
6.3.1. Индукция энзимов
Многие химические вещества, как эндогенные, так и поступающие из окружающей среды,
обладают способностью усиливать синтез в организме энзимов биотрансформации ксенобиотиков.
Этот феномен, получивший название индукции энзимов, существенным образом определяет
чувствительность живых существ к действию токсикантов. Несколько сот химических веществ
совершенно разного строения, как установлено, являются индукторами монооксигеназ и других
ферментативных систем. К числу сильных индукторов микросомальных ферментов принадлежат
многие лекарства и промышленные токсиканты. Все индукторы - жирорастворимые органические
вещества. Их действие, как правило, неспецифично, то есть индуктор вызывает повышение
активности более чем одного энзима. Индукция возможна, как правило, при повторном введении
соединения.
6.3.1.1. Индукторы метаболизма
Многочисленные индукторы монооксигеназных систем можно отнести к одному из двух классов.
Представителем первого класса является фенобарбитал, другие барбитураты, некоторые
лекарства и инсектициды. Ко второму классу индукторов относятся в основном полициклические
углеводороды: ТХДД, 3-метилхолантрен, бенз(а)пирен и т.д. Самым сильным из известных
индукторов монооксигеназ является 2,3,7,8-тетрахлордибензо-р-диоксин (ТХДД). Его эффективная
доза составляет 1 мкг/кг массы. В подавляющем большинстве случаев ксенобиотики проявляют
свойства индукторов, действуя в значительно больших дозах (более 10 мг/кг).
Фенобарбитал вызывает выраженную пролиферацию гладкого эндоплазматического ретикулума в
гепатоцитах и увеличение активности Р-450. В результате возрастает мощность таких процессов,
как деметилирование ксенобиотиков (нитроанизол), гидроксилирование (барбитураты),
эпоксидирование (альдрин).
Индукция, вызываемая полициклическими углеводородами не сопровождается пролиферацией
гладкого эндоплазматического ретикулума, но при этом существенно возрастает активность Р-450,
УДФГ-трансферазы, гидроксиолаз.

Некоторые индукторы способны специфично активировать отдельные изоформы Р-450. К числу
таковых относятся, в частности, прегненолол-16 -карбонитрил (ПКН), активирующий 3А1
изоформу Р-450, этанол, индуцирующий 1А2 изоформу Р-450, клофибрат - 4А изоформу.
Поскольку ксенобиотики, как правило, вызывают индукцию более чем одной ферментативной
системы (барбитураты, полигалогенированные бифенилы одновременно вызывают индукцию Р-
450, УДФГТ, GST и др.), предсказать влияние индукторов на токсикокинетику и токсикодинамику
ксенобиотика практически не возможно. Эффект может быть определен только экспериментально.
Задача усложняется еще и тем, что индукция того или иного энзима, вызванная разными
индукторами, не одинаково сказывается на скорости метаболизма разных ксенобиотиков (таблица
6).
Таблица 6. Влияние некоторых индукторов метаболизма на активность УДФ-
глюкуронилтрансферазы печени крыс (при использовании -нафтола, морфина и
хлорамфеникола в качестве субстратов)
Индукторы
Активность УДФ-ГТ (нмол/мин/мг белка)
-нафтол морфин хлорамфеникол
Контроль 75 +/- 15 7,9 +/- 0,6 0,36 +/- 0,1
Фенобарбитал (100 мг/кг/сут) 84 +/- 20 18,0 +/- 2,8 1,78 +/- 0,3
3-метилхолантрен (40 мг/кг) 212 +/- 49 9,9 +/- 1,7 0,39 +/- 0,1
Арохлор 1254 (80 мг/кг) 143 +/- 25 13,0 +/- 2,1 0,76 +/- 0,2
(K.W. Bock, 1977)
6.3.1.2. Механизмы индукции
Индукция предполагает синтез дополнительного количества того или иного энзима в органах и
тканях de novo. Ингибиторы синтеза белка (пуромицин, этионин, циклогексимид), а также
ингибиторы синтеза РНК (актиномицин Д) блокируют индукцию микросомальных энзимов.
Поскольку блокаторы синтеза ДНК (гидроксимочевина) не эффективны, можно сделать вывод, что
феномен индукции энзимов биотрансформации ксенобиотиков реализуется на уровне
транскрипции генетической информации.
Механизм феномена полностью не изучен. Одна из первых гипотез была предложена S. Granick в
1966 году для объяснения индукторных свойств стероидов. Автор полагал, что индукция
эндоплазматического ретикулума гепатоцитов, содержащего цитохромы, регулируется
концентрацией в клетке свободного гема, высвобождающегося в ходе обменных процессов в
эндоплазматическом ретикулуме. Гем взаимодействует с внутриядерным апорепрессором. При
этом образуется репрессор, который в свою очередь угнетает ген-оператор. В тот момент, когда
ген-оператор находится в неактивной форме, на соответствующем участке ДНК не
осуществляется синтез мРНК, необходимой для синтеза энзиматических белков, входящих в
структуру эндоплазматического ретикулума. При отсутствии свободного гема или в тех случаях,
когда гем не может присоединиться к апорепрессору, происходит активация синтеза мРНК и
выраженная пролиферация эндоплазматического ретикулума. Естественными ингибиторами
образования репрессора и являются стероиды, активно взаимодействующие с апорепрессором. В
соответствии с этой моделью около 300 ксенобиотиков (далеко не все структурные аналоги
стероидов) способны избирательно взаимодействовать с апорепрессором и блокировать тем
самым образование репрессора. Эти вещества проявляют свойства индукторов микросомальных
ферментов. Кроме того, так как большинство индукторов являются, как и стероиды,
жирорастворимыми веществами они активно захватываются мембранными структурами
цитоплазмы гепатоцитов, вытесняя при этом из соответствующих сайтов связывания
"депонированные" стероиды. Концентрация последних повышается в цитоплазме, а затем и в ядре
клеток. Связывание с апорепрессором усиливается, активируется процесс синтеза энзимов
метаболизма ксенобиотиков.
в соответствие с более поздними представлениями механизм действия стероидных гормонов,
полициклических ароматических углеводородов, 2,3,7,8,-тетрахлордибензо-п-диоксина, состоит во
взаимодействии с цитозольными рецепторными белками. Образующиеся комплексы мигрирует в
ядро клетки, где вызывают дерепрессию регуляторных генов и, тем самым, активирует синтез того
или иного энзима. В случае ТХДД такой рецепторный цитоплазматический протеин
идентифицирован, по крайней мере, в гепатоцитах лини мышей, чувствительных к ароматическим
углеводородам. Установлено, что синтез гидроксилазы ароматических улеводородов (aryl
hydrocarbon hydroxylase) регулируется локусом единственного доминантного гена, Ah, и может
быть усилен при введении ТХДД. Цитозольный белок-регулятор гена получил название Ah-
рецепторный протеин.
Другие индукторы, такие как барбитураты, вероятно действуют с помощью иного механизма. Хотя
известно, что в основе процесса также лежит индукция синтеза белка, до конца не выяснено каким
образом клетка распознает индуктор и как осуществляется воздействие на процесс транскрипции.
Индукторы класса фенобарбитала относятся к числу малоактивных соединений. Для реализации
эффекта нужны дозы препаратов на несколько порядков превышающие эффективные дозы ТХДД.
Рецепторный белок для фенобарбитала пока не идентифицирован.
Помимо усиления синтеза энзимов дополнительным механизмом индукции является стабилизация
информационной РНК и белковых молекул в клетке.
6.3.1.3. Влияние индукторов на токсичность ксенобиотиков
Достаточно часто усиление метаболизма ксенобиотиков приводит к снижению их токсичности. Так,
повторное введение фенобарбитала белым крысам самцам приводит к увеличению
резистентности животных примерно в полтора раза к таким высоко токсичным ФОС, как зарин,
зоман, ДФФ и др. Понижается чувствительность экспериментальных животных к цианидам. Вместе
с тем токсичность других веществ, при этом, существенно возрастает. Например, усиливается
гепаттоксическое действие алкалоида монокротолина и циклофосфамида, канцерогенная
активность 2-нафтиламина. Вследствие индукции усиливается также токсичность
четыреххлористого углерода, бромбензола, иприта и др.
Другим последствием индукции может быть изменение соотношения интенсивности метаболизма
ксенобиотиков в разных органах и тканях, в результате чего основным органом биопревращения
ксенобиотика у экспериментального животного, получавшего индукторы, становится иной орган,
чем у интактных животных. Так, после введения крысам 3-метилхолантрена (индуктор) основным
органом метаболизма 4-ипомеанола (токсичный дериват фурана) становятся не легкие (как в
норме), а печень.
Индукторы из группы производных барбитуровой кислоты способны одновременно активировать
синтез одних изоферментов (например, цитохромР-450 зависимых оксидаз) и угнетать активность
других. В этой связи a priori трудно предсказать последствия влияния индукторов на токсичность
ксенобиотиков.
У человека индукция микросомальных ферментов нередко становится следствием различных
привычек (курение, прием алкоголя и т.д.), профессионального и экологического контакта с
веществами (ПАУ, органические растворители, диоксины, галогенированные инсектициды и т.д.),
длительного приема некоторых лекарств (барбитураты, антибиотики типа рифампицин и т.д.).
6.3.2. Угнетение активности энзимов
Многие вещества способны угнетать активность ферментов, катализирующих метаболизм
ксенобиотиков.
Группа ингибиторов метаболизма включает:
- конкурентные ингибиторы ферментов (альтернативные субстраты). Например, этиловый спирт -
ингибитор метаболизма метанола или этиленгликоля; никотинамид - угнетает N-деметилирование
аминопирена и т.д.;
- неконкурентные ингибиторы. Это, как правило, алкилирующие агенты, угнетающие активность
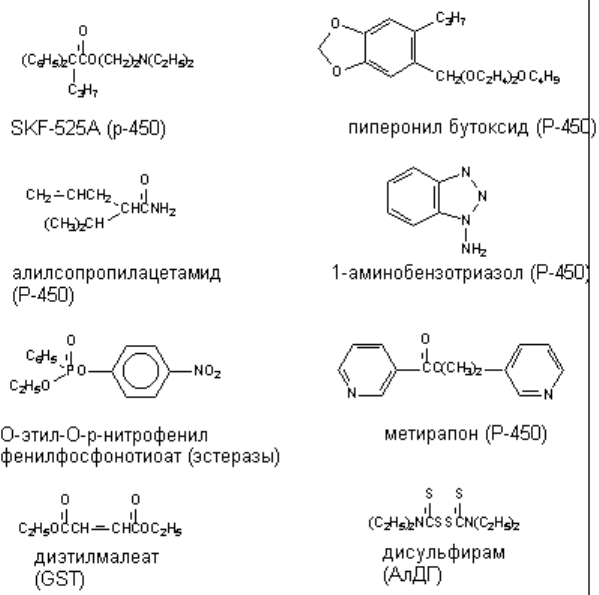
энзима, но не конкурирующие с субстратом. Например, метирапон является хорошо известным
ингибитором монооксигеназных реакций биопревращения. К этой же группе относится вещество
SKF-525 - известный ингибитор Р-450;
- "суицидные ингибиторы" - вещества, образующиеся в процессе метаболизма ксенобиотика при
участии данного фермента и одновременно являющиеся его ингибиторами. Например,
ингибиторами Р-450 такого рода являются дигидропиридины; метаболиты пиперонилбутоксида
угнетают микросомальное окисление многих ксенобиотиков в печени, таких как альдрин, анилин,
аминопирен, карбарил и др;
- реакционноспособные промежуточные метаболиты, ингибирующие активность энзимов
нескольких типов в месте их образования: метаболиты четыреххлористого углерода, дихлорэтана
и т.д.;
- ингибиторы синтеза кофакторов и простетических групп энзимов. К числу таких относятся,
например, Со, блокирующий синтез гема, являющегося простетической группой цитохромР-450
зависимых оксидаз; вещества истощающие запасы глутатиона в клетках.
Ингибиторы ферментов метаболизма не нашли в настоящее время практического применения.
Однако в условиях лаборатории для исследовательских целей некоторые из них используются
достаточно часто (рисунок 16).
Рисунок 16. Некоторые ингибиторы метаболизма ксенобиотиков. В скобках указаны ингибируемые
энзимы
Если ксенобиотик подвергается в организме детоксикации, угнетение его метаболизма приведет к
повышению токсичности, если происходит биоактивация, токсичность вещества понижается.
Например, дисульфирам (антабус), являясь ингибитором альдегиддегидрогеназы, вызывает
резкое повышение содержания уксусного альдегида в крови и тканях человека принявшего этанол.