Hugo W.B., Russel A.D.(ed). Pharmaceutical Microbiology
Подождите немного. Документ загружается.

Hammond S.M., Lambert P.A. & Rycroft A.N. (1984) The Bacterial Cell Surface. London: Croom Helm.
Hinkle P.C. & McCarty R.E. (1976) How cells make ATP. SciAm, 238, 104-123.
Nikaido H. & Vaara T. (1986) Molecular basis of bacterial outer membrane permeability. Microbiol
Rev, 49, 1-32.
Russell A.D. & Chopra I. (1996) Understanding Antibacterial Action and Resistance, 2nd edn.
Chichester: Ellis Horwood.
The references below refer to the subject matter in 5.6.
Microscopy, DEFT
Pettipher G.J., Mansell R., McKinnon C.H. & Cousins, CM. (1980) Rapid membrane filtration—
epifluorescent technique for direct inumeration of bacteria in raw milk. Appl Environ Microbiol,
39,423-429.
Denyer S.P. & Ward K.H. (1983) A rapid method for the detection of bacterial contaminants in
intravenous fluids using membrane filtration and epifluorescent microscopy. J Parental Sci Technol,
37, 156-158.
Flow cytometry
Shapiro H.M. (1990) Flow cytometry in laboratory microbiology: new directions. Am Soc Microbiol
News, 56, 584-586.
Microcalorimetry
Beezer A.E. (1980) Biological Microcalorimetry. London: Academic Press.
Impedance
Silley P. & Forsythe S. (1996) Impedance microbiology—a rapid change for microbiologists. J Bacterial
80, 233-243.
Bioluminescence
Stanley P.E., McCarthy B.J. & Smither R. (eds) (1989) ATP Luminescence: Rapid Methods in
Microbiology. Society of Applied Bacteriology Technical Series No. 26. Oxford: Blackwell Scientific
Publications.
Stewart G.S.A.B., Loessner M.J. & Scherer S. (1996) The bacterial lux gene bioluminescent biosensor
revisited. Am Soc Microbiol News, 62, 297-301.
General reference
Stannard C.J., Petit S.B. & Skinner F.A. (1989) Rapid Microbiological Methods for Foods, Beverages
and Pharmaceuticals. Society of Applied Bacteriology Technical Series No. 25. Oxford: Blackwell
Scientific Publications.
Yeasts and moulds
1 Introduction 4 Cryptococcus neoformans
2 Saccharomyces cerevisiae 5 Neurospora crassa
2.1 The life cycle
2.2 Metabolism and physiology 6 Penicillium and Aspergillus
2.3 Cell wall
7 Epidermophyton, Microsporum and
3 Candida albicans Trichophyton
3.1 Pharmaceutical and clinical significance
3.2 Alternative morphologies 8 References
Introduction
Yeasts and moulds are members of the fungi. Yeasts are characterized as being essentially
unicellular, whereas moulds are composed of filaments which en masse frequently
appear fuzzy or powdery. The familar budding yeast Saccharomyces cerevisiae, also
known as Baker's or Brewer's yeast, is usually thought of as the typical yeast. The
green mould Penicillium digitatum, a frequent spoiler of fruits such as apples or oranges,
and the bread mould Neurospora crassa will also be well-known to many. These latter
two organisms are properly considered as typical moulds. As is usually the case, however,
life is not completely straightforward for there are a considerable number of so-called
'dimorphic fungi' which can alternate between yeast-like and filamentous forms. One
such organisms is Candida albicans. To make matters more complicated, it has been
rediscovered that the would-be typical yeast S. cerevisiae can also form filaments
under a variety of different conditions (Gimeno et al. 1992). All of these fungi have
pharmaceutical and medical significance. The precise nature of this significance is
different in each case. For example, S. cerevisiae is generally regarded as a totally safe
organism suitable for use in human food and drink; the reason for its importance is
because it is by far the best understood eukaryotic organism on the planet. In contrast,
Cryptococcus neoformans has a variety of ways by which it can evade defence
mechanisms of the immune system, but is relatively little studied. In between these
two extremes are many yeasts and moulds, which are omnipresent in the environment,
in or on our foods, or a part of the normal flora of humans, but all of which can
opportunistically contaminate pharmaceutical preparations or cause post-operative
disease. All fungi pose a threat to immunocompromised individuals. This knowledge
should be weighed against a background of a general lack of suitable antifungal agents
(see Chapter 5). The approach of this chapter will be to first describe S. cerevisiae in
considerable detail because so much is known about it. Then, other yeasts and moulds
will be considered in turn, pointing out (where appropriate) significant differences
from S. cerevisiae or from each other.
Yeasts and moulds 35
2
Saccharomyces cerevisiae
Saccharomyces cerevisiae has a predominant place in the realms of cell biology and
molecular biology where it has become accepted as the universal model eukaryote.
The main reason for this is its genetic tractability. Traditionally, for reasons associated
with its importance to the food and drink industry, a great deal was known about the
biochemistry and physiology of this yeast. Later, with the advent of yeast genetics, a
vast range of well-characterized mutants became available. In turn, because S. cerevisiae
can be transformed and is readily amenable to genetic manipulation, this permitted
the isolation and characterization of many yeast genes. Ultimately, in mid 1996
the nucleotide sequence of the entire genome of the organism was reported. This
achievement is still only a far-off dream for molecular biologists studying most other
eukaryotic organisms. Nevertheless, it is possible to identify genes from other organisms
by means of genetic complementation in S. cerevisiae. Explained briefly, only one
piece of DNA from another organism will be able to substitute for a mutation in a
known gene in S. cerevisiae—this is a segment of DNA which carries the homologous
gene (i.e. codes for the same function) in the other organism. The availability of well-
defined mutants in S. cerevisiae combined with the facility of genetic manipulation
and this yeast's short generation time, make this a very rapid way to identify heterologous
(i.e. belonging to another organism) genes. Many of the latest concepts in cell and
molecular biology (e.g. concerning control of the cell cycle) have been developed and
tested in this organism. Naturally then, since it is the prime model eukaryote, it is also
the best understood fungus.
The life cycle
The life cycle of S. cerevisiae is shown in Fig. 2.1. It can exist both as a haploid (one
copy of each chromosome per cell) or as a diploid (two copies of each chromosome per
cell). Haploids exists as one of two sexes referred to as mating type a and mating type
a. When two haploid cells come close together they cause each other to arrest in the
Gl phase of the cell cycle. Each subsequently produces a special protuberance enabling
growth towards the mating partner. These somewhat abnormal looking cells are termed
'schmoos'. A haploid will only mate with another haploid of the opposite mating type.
This is achieved by the expression of specific oligopeptide mating pheromones
(hormones with brings about behavioual change in cells of the opposite sex) and the
possession of surface receptors only for the opposite pheromone (hence, mating type a
strains produce only a-factor and have receptors for a-factor, whilst mating type a
strains produce only cu-factor and have receptors for a-factor). The resulting diploid,
like the haploids from which it arose, is capable of repeated rounds of vegetative
reproduction.
The vegetative cell cycle of S. cerevisiae has received extensive attention. There
are many justifications for this. Firstly, the cell cycle in this organism has many
convenient 'landmarks' (Hartwell 1974, 1978; Pringle 1978) which make it very easy
to identify the exact point in the cell cycle at which a cell happens to be. Examples of
these landmark events include bud emergence, the size of the bud, mitosis (nuclear
division takes place through the neck between the 'mother' cell and the bud), and cell
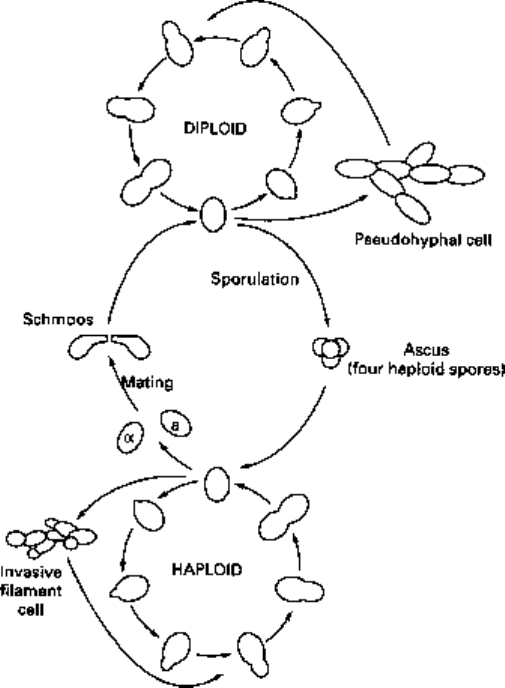
Fig. 2.1 The life cycle of Saccharomyces cerevisiae.
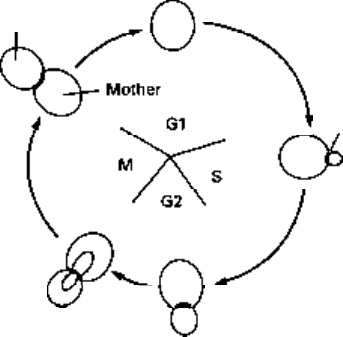
separation. Other markers of cell cycle progress are also apparent to the more experienced
observer (Fig. 2.2). The reader will notice from Fig. 2.2 that the 'daughter' which is
formed is smaller than the mother cell from which it arose. There is a size control
which operates over initiation of anew cell cycle (Pringle & Hartwell 1981), and since
the mother is larger than the minimum size necessary to pass this control, but the daughter
is not, the consequence is that the mother cell can immediately start a new cell cycle,
whereas the daughter must first grow for a period until it is large enough. Hence, mother
and daughter do not proceed through the next cell cycle synchronously. The significant
extent of morphological change throughout the cell cycle provides another reason for
studying this yeast as the construction of defined morphology.
A third justification for studying the cell cycle of this yeast is that it affords
a convenient system in which to study cell polarity. Together with asymmetric cell
division (inherent in the S. cerevisiae cell cycle with the unequal sized mothers and
daughters), the development of polarity is crucial in many aspects of development and
differentiation. Furthermore, as explained in more detail later in this chapter, the correct
Yeasts and moulds 37
Daughter
Cell
separation
Unbudded cell
Nuclear
division
Chitin
ring
Bud
emergence
Bud
enlargement
Fig. 2.2 'Landmark' events in
the cell cycle of Saccharomyces
cerevisiae. Gl, S, G2 and M are
the classical phases of the
eukaryotic cell cycle.
development of polarity is an essential aspect in the life of most fungi. The development
of polarity and the resulting asymmetric division can be considered as five constituent
processes (Lew & Reed 1995) shown schematically in Fig. 2.3. These are:
1 the F-actin cytoskeleton;
2 the polarity of growth achieved by the way new cell wall material is arranged;
3 the location of 10-nm neck filaments;
4 formation of the cell 'cap';
5 the distribution of DNA and microtubules.
The construction of a yeast cell requires isotropic growth. Bud emergence is signalled
by the accumulation of secretory vesicles, the rho protein Cdc42p and a cap of
membrane-localized actin patches. Once the cap is established, subsequent bud
emergence is accomplished entirely by polarized growth. Following bud emergence,
the rings of 10-nm filaments remain at the mother/bud neck, whereas the proteins in
the cap concentrate at the tip of the bud where secretion takes place. Later, there is a
critical switch back to isotropic growth which brings about swelling of the bud. Most
of the proteins of the cap appear to disperse simultaneously with the apical/isotropic
switch. At cytokinesis, secretion is redirected to the neck and the proteins of the cap
redistribute to this region. Mutants have been isolated which distinguish between the
separate components and processes. In turn, the genes which the mutations have
identified have all been characterized.
The pattern of budding in haploids differs from that in diploids (Friefelder 1960).
Haploids grown in rich medium bud in an axial pattern, i.e. each new bud site is placed
adjacent to the previous one. In the same rich nutrient conditions diploids exhibit bipolar
budding, in this case choosing new bud sites at either end of the cell (Fig. 2.4). Under
a variety of other conditions, all presumably involving some form of nutrient limitation,
diploids will form pseudohyphae and haploids will form invasive filaments. As alluded
to earlier in this chapter, this represents a 'rediscovery' in the case of S. cerevisiae
because it had been known for a long time and forms part of the basic taxonomy. Its
significance had been ignored. This situation arose because the ability to form these
structures had been crossed-out of the genetic background of many academic strains
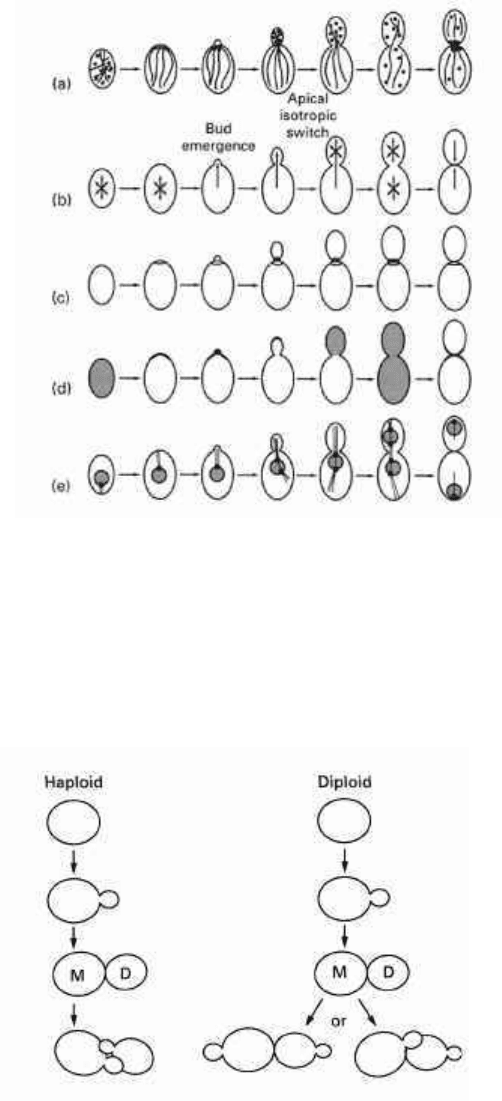
Fig. 2.3 The development of polarity and asymmetric division in Saccharomyces cerevisiae. The
diagram is reproduced in a slightly simplified form from the work of Lew & Reed (1995) with the
permission of Current Opinion in Genetics and Development, (a) The F-actin cytoskeleton: strands =
actin cables; (•) cortical actin patches, (b) The polarity of growth is indicated by the direction of the
arrows; (arrows in many directions signifies isotropic growth), (c) 10-nm filaments which are
assembled to form a ring at the neck between mother and bud. (d) Construction of the 'cap' at the
pre-bud site. Notice that the proteins of the cap become dispersed at the apical/isotropic switch, first
over the whole surface of the bud, then more widely. Finally, secretion becomes refocussed at the neck
in time for cytokinesis, (e) The status and distribution of the nucleus and microtubules of the spindle.
Notice how the spindle pole body (•) plays an important part in orientation of the mitotic spindle.
Fig. 2.4 The budding pattern in
haploid and diploid
Saccharomyces cerevisiae. The
original cell which formed a bud
is the mother (M). The daughter
cell (D) is shown remaining
attached as might be the case in
i colonies growing on the surface
of agar.
around the world. Pseudohyphae are chains of regular-shaped, elongated cells in which
unipolar budding predominates. The analogous situation in colonies of haploids growing
on solid media is the formation of invasive filaments which are capable of penetrating
Yeasts and moulds 39
the agar. The generally accepted view is that starvation of nitrogen is the signal for the
switch from the yeast to a filamentous form (Kron et al. 1994), although it has also
been shown that pseudohyphal growth is strictly oxygen-dependent (Wright et al. 1993)
and that limitation of oxygen during continuous cultivation can result in the formation
of pseudohyphae (Kuriyama & Slaughter 1995). In 1996 Dickinson showed that,
dependent upon the concentration used, 'fusel' alcohols, i.e. n-amyl, isoamyl alcohol,
etc., caused the formation of hyphal-like extensions or pseudohyphae in a wide number
of different yeast species which were being cultured in rich liquid media where the
cells would normally proliferate as yeasts rather than in any other form (Dickinson
1996). It seems reasonable to conclude that since fusel alcohols are produced when the
yeasts are under various conditions of nutrient stress, the many situations which have
been reported to induce pseudohyphal formation are triggered by fusel alcohols. As we
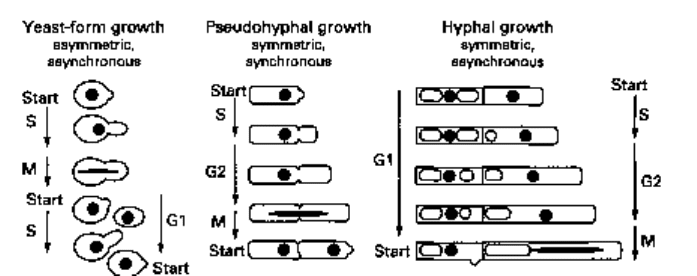
have already noted, yeast-form proliferation is asymmetric and asynchronous; in
contrast, as others have already observed (Kron & Gow 1995), pseudohyphal growth
is symmetric and synchronous and, as will become apparent later in this chapter, hyphal
growth is symmetric and asynchronous (Fig. 2.5).
The diplophase and haplophase are equally stable. Hence, in the presence of adequate
nutrients, both are capable of repeated rounds of vegetative growth and mitosis.
However, in the presence of a poorly utilized carbon source such as acetate, and usually
in the absence of a nitrogen source, diploid strains switch to the alternative developmental
pathway of meiosis and spore formation. This process of sporulation gives rise to
structures termed 'asci'. Each single ascus contains four haploid ascospores (usually
referred to simply as 'spores'). Sporulation in diploid strains of S. cerevisiae has been
studied as a simple unicellular model of differentiation because it involves the co-
ordination of a complex sequence of genetic, biochemical and morphological events
(Fig. 2.6). The developmental switch occurs only in the Gl phase of the cell cycle, in
normal (a/a) diploids which are respiratorily complete. Hence, it requires the co-
ordination of signals about the environment, about the physiological and metabolic
Fig. 2.5 Cell cycles resulting in yeast-form cells, pseudohyphae and hyphae. In many respects the
cell cycle of pseudohyphal cells is similar to that of yeast-form cells, except that in pseudohyphae
G2 is prolonged, thus larger daughter cells are produced which are identical in size to the mother
cell. Hence, mother and daughter are both sufficiently large to start the next cell cycle and so bud
synchronously. In hyphae the apical cell becomes progressively longer. The diagram is reproduced
from the review of Kron & Gow (1995) with the permission of Current Opinion in Cell Biology.
40 Chapter 2
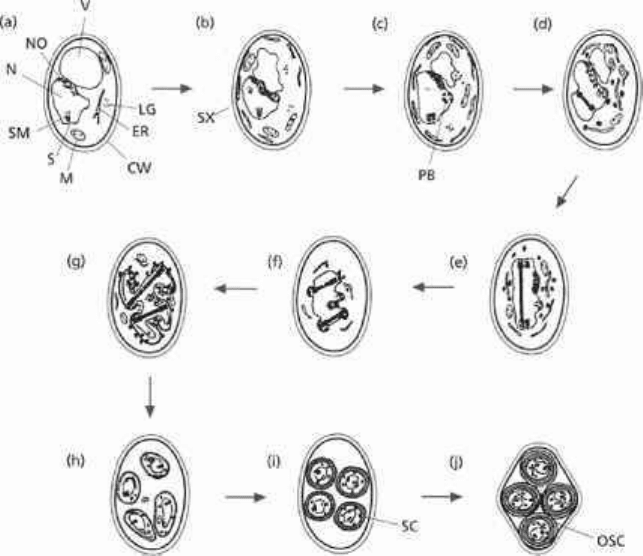
Fig. 2.6 The morphological events of sporulation in Saccharomyces cerevisiae. (a) starved cell: V,
vacuole; LG, lipid granule; ER, endoplasmic reticulum; CW, cell wall; M, mitochondrion; S, spindle
pole; SM, spindle microtubules; N, nucleus; NO, nucleolus, (b) Synaptonemal complex (SX) and
development of polycomplex body (PB) along with division of spindle pole body in (c). (d) First
meiotic division which is completed in (e). (f) Prepararation for meiosis II. (g) Enlargement of
prospore wall, culminating in enclosure of separate haploid nuclei (h). (i) Spore coat (SC) materials
produced and deposited, giving rise to the distinct outer spore coat (OSC) seen in the completed
spores of the mature ascus (j). Reproduced from the review by Dickinson (1988) with permission
from Blackwell Science Ltd.
status of the cell along with a way of monitoring the cell's ploidy and position in
the cell cycle. It is attractive for study because it involves meiosis, a relatively rare
event that occurs only in cells of specialized tissues in higher eukaryotes, and because
it allows the study of developmentally regulated gene expression. Transfer to the
sporulation pathway also involves a number of distinct metabolic switches (Dickinson
1988; Dickinson & Hewlins 1991). The whole process can be completed within 24
hours. The products of sporulation (haploid ascospores) have far greater resistance to
heat, solvents, dehydration, etc. than vegetative cells. If returned to good nutrient
conditions the spores will germinate and commence proliferation as free-living haploids.
This whole sequence of events forms the basis of conventional genetics and laboratory
strain construction in this yeast. Two haploids of opposite mating type each carrying
particular mutations are placed in close proximity to each other on the surface of an
agar medium. After mating and subsequent formation of the diploid, the cells can be
replica plated to a different medium to allow selection for the diploid and against the
parental haploids. ('Replica plating' involves making a replica of a group of cells which
Yeasts and moulds 41
are growing on one type of medium onto one, or more, different media. This is done by
pressing the agar surface of a Petri dish carrying cells onto a sheet of sterile velvet.
Subsequently, other, uninoculated, Petri dishes can receive doses of these cells by being
pressed onto the surface of the velvet). This is most simply arranged by ensuring that
each parental haploid has different auxotrophic requirements, hence the resultant diploid
will be able to grow on a minimal medium (due to complementation) whereas neither
of the parents can. After 1-3 days growth, the diploid will then be replica plated again
onto a sporulation medium. When the asci have formed, the individual spore progeny
can be separately grown as individual clones (a clone is a group of cells which are
genetically identical). This final step is accomplished by enzymatic digestion of the
ascus wall followed by micromanipulation of the individual spores (a process known
as 'dissection'). Due to the fact that meiotic recombination took place during sporulation,
the spores will have different combinations of mutations to those present in the original
parents. The precise combination of mutations in each spore can be determined by
analysing the phenotypes.
Metabolism and physiology
Saccharomyces cerevisiae is normally described as a faculative anaerobe which means
that it is able to proliferate under either anaerobic or aerobic conditions. It is able to
utilize a wide range of mono-, di- and oligosaccharides, ethanol, acetate, glycerol,
pyruvate and lactate. The favourite carbon source is glucose and the preferred mode of
metabolism is fermentative using the Embden-Meyerhof pathway (EMP) resulting in
the formation of ethanol. Many aspects of metabolism and physiology in this organism
(not merely carbon metabolism) are subject to catabolite repression which in most
cases means glucose repression. In the presence of glucose, synthesis of the enzymes
necessary for disaccharide (sucrose and maltose) or galactose utilization and for growth
on non-fermentable carbon sources (ethanol, acetate, glycerol, pyruvate and lactate) as
well as mitochondrial development are repressed. As the repressing substrate (glucose)
is consumed its concentration falls and the cells are said to become 'derepressed'; this
occurs typically at glucose concentrations below 0.2%. In other words, induction of
respiratory enzymes and components of the mitochondrial electron transport chain
occurs. This metabolic switch takes place late in the exponential phase of a batch
culture. As the cells pass through the deceleration phase and enter the stationary
phase they will be fully derepressed and will start to consume the ethanol that was
produced earlier. This requires the full participation of the tricarboxylic acid (TCA)
and glyoxylate cycles for the complete oxidation of ethanol to carbon dioxide and
water. Cells utilizing any of the non-fermentable carbon sources are also carrying out
gluconeogenesis. The glucose-6-phosphate produced as a result of this gluconeogenesis
is used both for the production of storage carbohydrate (trehalose) and for 'shuttling'
around the hexose monophosphate pathway (HMP) for synthesis of ribose which is
required for nucleotide (and hence ultimately nucleic acid) biosynthesis. The importance
of the glycolytic pathway to S. cerevisiae cannot be overstated. This is underlined by
the frequently quoted figure that the enzymes of glycolysis represent 30-65% (depending
upon physiological conditions) of soluble protein (Fraenkel 1982). The storage material
trehalose is produced in large quantities during sporulation (Dickinson et al. 1983). It
confers to the spore the ability to withstand dehydration. A wide range of organisms
in low water environments utilize trehalose for the same purpose including most
insects and the remarkable drought-resisting resurrection plant Selaginella lepidophylla
which can survive protracted desiccation until rains return (Leopold 1986). Trehalose
does this by preventing phase transitions within membranes (Crowe et al. 1984). The
compound is now added to a considerable number of laboratory products in order to
extend their shelf-life and its use in pharmaceutical and medical materials including
plasma, blood-based products, whole cells and tissues is under active investigation for
the same reason.
Notwithstanding the foregoing, an important constraint on this otherwise meta-
bolically flexible organism is the fact that proliferation under truly anaerobic conditions
(something that is very difficult to achieve in the laboratory) is not possible without the
provision of unsaturated fatty acid and sterol (Andreasen & Stier 1953, 1954). These
are required for the assembly of membranes. Naturally, mutants defective in fatty acid
or sterol biosynthesis have such requirements, but so do mutants with defects in
porphoryin biosynthesis due to the involvement of haematin in the synthesis of both
groups of compounds. Wild-type S. cerevisiae do not take up sterol under aerobic
conditions. It is possible to supply a limited range of alternative sterols instead of
the yeast's usual ergosterol. The ability of such an alternative sterol to support growth
of anaerobic S. cerevisiae is a way of assessing the structural specificity of sterol
requirement (Henry 1982). Some yeast sterol mutants were isolated as auxotrophs
requiring ergosterol whilst others were obtained on the basis of resistance to the polyene
antibiotic nystatin. Polyene antibiotics alter membrane permeability by interaction
with specific membrane sterols (Cass et al. 1970; Norman et al. 1972; see Chapter 8)
and seem not to inhibit lipid synthesis. Hence, mutants resistant to polyene antibiotics
have been useful in identifying the effects of altered sterol composition on different
membranes within the cell. This can be reflected in, for example, altered permeability
to a specific molecule or ion.
2.3 Cell wall
The cell wall of S. cerevisiae, like that of other fungi, is very strong. Despite its great
strength, one should remember that the cell wall is a dynamic structure (unlike a brick
wall). There are three major components:
1 an internal glucan layer,
2 the external layer of mannoproteins;
3 chitin which occupies various specialized locations.
Cell wall composition varies according to physiological conditions and developmental
status. For example, the wall of cells from stationary phase is much more resistant to
degradation by /3-glucanase than that from exponential phase cells (Necas 1971). The
glucan of S. cerevisiae is mainly j8(l-3)-linked glucoses with branching via /?(l-6)-
linked glucose units (Manners et al. 1973a, b). Most of the mannoproteins can only be
released after enzymatic degradation of the glucan layer. There are long «(l-6)-
mannose chains with a{\-2) and cu(l-3) side chains iV-linked to asparagine. There are
also short mannose chains O-linked to serine or threonine (Van Rinsum et al. 1991).
The carbohydrate chains of the mannoprotein layer are the main antigenic determinants
Yeasts and moulds 43