Heitjans P., Karger J. (Eds.). Diffusion in Condensed Matter: Methods, Materials, Models
Подождите немного. Документ загружается.

12 Diffusion in Membranes 477
The fluid-like nature of membranes is the key to their various dynamic
properties. Out-of-plane fluctuations such as undulations [24] are more like a
rule than exception, since their activation energy is of the order of few k
B
T ’s.
Fluid-like lipid bilayers have no resistance to shear, unless the cytoskeleton is
attached to the membrane. As the molecules are able to pass one another, the
lateral diffusion coefficients of individual lipid molecules are relatively high,
typically of the order of 10
−10
–10
−12
m
2
/s. Then, as the lipids and proteins
are complex chain-like molecules, they have various internal degrees of free-
dom, and thus rotational diffusion characterizing their rotation rates around
the symmetry axes is also important. The decay of density fluctuations, in
turn, gives rise to the so-called collective diffusion (related to area fluctua-
tions of the bilayer), while the diffusive transport of lipids across membranes
yields another interesting transport process known as the flip-flop.
The lipid-bilayer component of membranes displays a number of different
phases separated by phase transitions. In the present context the most impor-
tant transition is the so-called main transition, which takes the bilayer from a
low-temperature solid phase to a high-temperature liquid (fluid) phase. The
solid phase has a two-dimensional crystalline character and in addition the
lipid acyl chains are conformationally ordered. This phase is therefore usu-
ally referred to as the solid-ordered phase. In the fluid phase, the crystalline
structure is lost and at the same time the acyl chains display a substan-
tial degree of conformational disorder. This phase is consequently termed
the liquid-disordered phase. The diffusional characteristics of the membrane
molecules are very different in the two phases.
Our aim in the next few sections is to provide the reader with some flavor
of the various diffusion processes taking place in membranes. Rather than
presenting an exhaustive description of the topic, we prefer a down-to-earth
approach on an introductory level, combined with instructive examples and
highlights of a few recent studies that we consider interesting.
12.3 Lateral Diffusion of Single Molecules
Here we discuss the motion of individual molecules in membranes as the
particles diffuse laterally in the bilayer.
12.3.1 Lateral Tracer Diffusion Coefficient
Let us consider a membrane comprised of two lipid monolayers (see Fig. 12.2).
Additionally, we may imagine that there are proteins embedded in the mem-
brane, and all these molecules are allowed to move in the plane of the mem-
brane. For simplicity’s sake the membrane is assumed to be flat (zero curva-
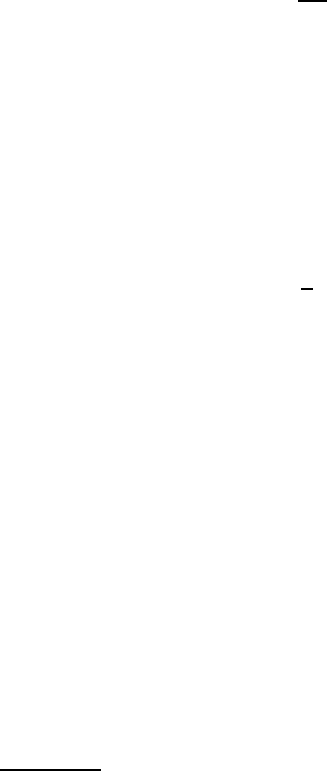
478 Ilpo Vattulainen and Ole G. Mouritsen
ture).
2
Furthermore, we can safely assume that the number of lipid molecules
in both monolayers remains constant, since the time scale of lipids exchang-
ing their positions from one monolayer to another (flip-flop) is of the order
of several seconds or even hours, while the lateral diffusion of lipids usually
takes place on much smaller time scales, typically nanoseconds.
A measure of how fast individual molecules move around is given by the
lateral tracer diffusion coefficient [27,28] (cf., e. g., also Chap. 10, (10.8) and
Chap. 18, (18.21))
D
T
= lim
t→∞
1
2dt
|r(t) − r(0)|
2
, (12.1)
where r(t) is the position of the tagged particle (or its center of mass) at time
t,andd = 2 is the dimensionality of a surface.
3
This definition is identical
to the original one by Einstein for the motion of Brownian particles in a
solvent [27].
We note that D
T
is essentially defined as a long-time limit of |r(t)|
2
,
which is the mean square displacement of a given particle. The tracer diffusion
coefficient, however, arises from correlations between consecutive displace-
ments of the particle at small times, as is demonstrated by the Green-Kubo
equation [28] (cf. Chap. 23, (23.15))
D
T
=
1
d
∞
0
dtφ(t), (12.2)
where φ(t) ≡v(t) · v(0) is the velocity correlation function of the tagged
particle (or its center of mass) in terms of its velocity v(t)attimet.
It can be shown that (12.1) and (12.2) are identical and yield the same
diffusion coefficient [28]. However, they probe slightly different properties of
the dynamics of the particle and therefore complement each other. While
the approach based on the mean square displacement assumes that one has
found the long-time limit (the diffusive regime) in which |r(t)|
2
∼t,the
Green-Kubo equation concentrates on the short-time regime and integrates
over velocity correlations prior to the long-time regime. At long times, beyond
some characteristic time scale τ
c
, the velocity correlation function φ(t)=0
for t>τ
c
.Itisobviousthatτ
c
is closely related to the onset of the diffusive
regime in which |r(t)|
2
∼t (cf, e. g., Chap. 21, Fig. 21.7).
In practice, the velocity correlation function provides a useful quantity
for examining the short-time dynamics through molecular dynamics simu-
lations, thus yielding valuable information of the microscopic details of the
diffusion process. Einstein equation, on the other hand, is probably more
useful as a means to determine the actual value of the diffusion coefficient
D
T
. Besides this, the mean square displacement provides one with a reliable
2
In the case of liposomes, the calculation for the diffusion coefficient has to be de-
termined in a spherical geometry. A further complication arises when the bilayer
is so soft that the diffusional motion changes the local curvature.
3
In the following discussion we assume that r(0) = 0.

12 Diffusion in Membranes 479
way to find the diffusive regime as well as to obtain information of effects
leading to anomalous diffusion (cf. Chap. 10). To illustrate these points, let
us argue that |r(t)|
2
∼t
x
,wherex is some time-dependent exponent. For
normal diffusion, x → 1 at long times, which allows one to find the regime
in which the Einstein equation is valid. However, there are also cases where
the heterogeneity of the membrane or large obstacles such as proteins hin-
der or bias the diffusion process in such a fashion that x<1evenatvery
long times [29]. Then the diffusion process is said to be subdiffusive (see
Sect. 12.3.3). The concept of anomalous diffusion is generally related to the
so-called Levy processes, which are discussed in more detail in [30, 31].
The diffusion of lipids and proteins in a bilayer is essentially a two-
dimensional process, and since we are dealing with a hydrodynamic medium
in which interparticle interactions are mediated by the solvent, it is worth-
while to point out the well-known problem of diffusion in two spatial di-
mensions. Under hydrodynamic conditions, φ(t) decays at long times as
φ(t) ∼ t
−d/2
[32, 33], and therefore for a truly two-dimensional system
φ(t) ∼ t
−1
. This seems to imply that (12.2) does not converge and thus
the diffusion coefficient is not well defined. However, one should keep in mind
that we are looking at the motion of lipids in a bilayer which is interacting
with the neighboring environment. These interactions imply that the mo-
mentum and the energy of the bilayer are not conserved. Thus, the motion
of lipids in a bilayer is a dissipative process, and the theoretical result above
does not apply in the present case. In brief, as long as anomalous diffusion
is not expected, one should find |r(t)|
2
∼t at long times with a constant
slope.
The two approaches mentioned above [(12.1) and (12.2)] are not unique,
although they provide the grounds since they can be considered as definitions
of the tracer diffusion coefficient. Of the other approaches suggested recently,
let us mention the method by which the calculation of D
T
can be optimized
by a so-called “memory expansion” of particle displacements [34]. Finally,
once D
T
has been found, one can use it to estimate the diffusion length
D
≡
√
Dτ, which the particle undergoes during a time interval of τ.
12.3.2 Methods to Examine Lateral Tracer Diffusion
To start with, we would like to emphasize the special nature of tracer diffusion
that should be accounted for in any study of diffusion. The tracer diffusion
process refers to a case where one follows the motion of some tagged individual
particle.
This is relatively easy in computer simulations of model systems, where
one can keep track of the positions and velocities of all particles one by
one. In experiments the situation is not as simple, however. Many techniques
are based on labeling lipid molecules by attaching rather bulky molecules
such as pyrene to their acyl chains or a colloidal particle to the head group.
The attached molecule has typically some property that allows one to follow
480 Ilpo Vattulainen and Ole G. Mouritsen
its position, fluorescent labeling being probably the most commonly used
technique. In some cases the fluorescence signal from a single lipid molecule or
lipid analogue yields a signal large enough to be detected on the background
of the noise. Most often one labels a certain fraction (such as 1/1000) of all
lipids, however. This leads to two possible problems. First, since the size of the
functional group attached to a parent molecule can be quite substantial, and
its diffusion characteristics may be very different from those of non-labeled
parent molecules, there is an obvious difficulty in interpreting experimental
diffusion data. Second, the fact that there are numerous labeled molecules in
a bilayer makes it possible that the trajectories of individual lipid molecules
cannot be followed any further. Consequently, one cannot use the Einstein
equation either. Nevertheless, as discussed later in this chapter, there are
certain ways to avoid this problem.
Computational Approaches
For studies of soft matter systems, the first-principles approach including
electronic degrees of freedom feels like a method of choice. However, its com-
putational costs are harsh and imply that it is limited to very small systems
over short time scales, and is therefore not applicable to membrane systems
as such. Current approaches of combining the first-principles techniques with
classical molecular dynamics (MD) may provide some relief to this prob-
lem [35, 36].
The classical MD approach ([37, 38], see also Chap. 23), however, has de-
veloped in recent years to a versatile method for studies of lipid and protein
dynamics in lipid bilayers [39–44]. In this approach, all atoms are treated
classically, their interactions are well defined, and the time evolution of par-
ticle positions is determined by solving Newton’s equations of motion. This
approach provides one with a classical but detailed description of the system,
and can yield plenty of relevant information of the structure and dynamics
of membranes at the atomic level. Nevertheless, the price one has to pay is
the computational cost. At present, state-of-the-art MD simulations are for
about 1000 lipid molecules in a bilayer plus a few thousand water molecules,
which allows one to study the system over a time scale of about 100 ns [45].
However, is this enough for studies of lateral lipid diffusion? To address this
question, let us first approximate the onset of the diffusive regime τ
c
using
aruleofthumbτ
c
≈ R
2
L
/D
T
,whereR
2
L
is the area per lipid in the plane of
the membrane and D
T
is its tracer diffusion coefficient. Using typical values
of R
L
=8
˚
AandD
T
=3×10
−11
m
2
/s for a single-component lipid bilayer of
DPPC molecules in a liquid-disordered phase [3], we obtain τ
c
≈ 20 ×10
−9
s.
Thus we have to look at times larger than about 20 ns to make sure that an
individual lipid molecule in a pure bilayer moves over a distance that is equal
to its own size. This time scale should be considered as the lower limit, since
displacements of this size can only barely be regarded as macroscopic ones.
12 Diffusion in Membranes 481
In particular in multicomponent systems often characterized by domain for-
mation, the time scale should be far larger since then R
L
needs to be replaced
by the characteristic size of the domains, which is typically 10 – 1000
˚
A [26].
We can conclude that detailed MD simulations are not the most feasible
approach of studying lateral diffusion. Consequently, less detailed approaches
have been suggested. The underlying idea is to simplify the model by replac-
ing an atomistic description with a coarse grained one that retains only some
of the most essential molecular features (see Fig. 12.1). In practice, this means
that the “fast” variables in a system are replaced with stochastic noise and
the interactions are chosen to represent those between coarse grained particles
rather than actual atoms. Recent progress on the field is encouraging since
some of the structural [46, 47] as well as dynamic [48, 49] properties of the
membrane have been successfully explored using this idea. However, one can
coarse grain the molecular description even further and describe the system
in terms of a minimal model, in which lipids and proteins are described in a
very simplified fashion in terms of disks or cylinders, for example. This ap-
proach allows studies of very large time and length scales and may be highly
useful for investigations of processes that are too complicated to be studied
by models including molecular information. Studies have shown [50–52] that
the approach of using minimal models works fine in a number of cases.
Experimental Techniques
The number of techniques which probe lateral tracer diffusion directly is lim-
ited. One popular technique is single-particle tracking [53, 54] in which one
attaches a colloidal particle (typically 40 nm in diameter) to the lipid or pro-
tein molecules by proper anti-body functionalization. Then, one follows the
trajectories of individual labeled molecules in bilayers by computer-enhanced
video microscopy, and statistical analysis of a large number of traces allows
one to determine the lateral tracer diffusion coefficient in terms of (12.1).
In recent years single-molecule fluorescence labeling assays have been devel-
oped which by ultra-sensitive fluorescence microscopy or fluorescence cor-
relation spectroscopy have permitted traces of single-molecule diffusion to
be observed [55–59]. An example of such observations is given in Fig. 12.4
which shows the motion of an individual fluorescence-labeled molecule in a
phospholipid bilayer and the recording of a part of a diffusion trace. The
time resolution of such experiments is in the range of 5 ms and the positional
accuracy is around 50 nm [58].
In principle, all other approaches we are aware of are based on follow-
ing some collective property of the whole system, such as the decay rate
of density fluctuations. As we will see in Sect. 12.5, this may lead to stud-
ies of the collective aspects of diffusion rather than tracer diffusion. Never-
theless, a number of techniques such as fluorescence recovery after photo-
bleaching (FRAP) [60, 61] (cf. also Chap. 16), nuclear magnetic resonance
(NMR) [62] (cf. also Chap. 10), (incoherent) quasi-elastic neutron scattering
482 Ilpo Vattulainen and Ole G. Mouritsen
(a)
(b)
Fig. 12.4. (a) Fluorescent image of a single fluorescence-labeled lipid molecule in a
phospholipid bilayer, and (b) the recording of a part of a diffusion trace of a single
lipid molecule. From [58].
(QENS) [63–66] (cf. also Chap. 13), and electron spin resonance (ESR) [67]
have been used to obtain information of single-molecule motion in mem-
branes. The time and length scales probed by these techniques differ sub-
stantially, for which reason they are often characterized as either microscopic
or macroscopic methods (see also Chaps. 1, 9 and, e. g., [68]). We will come
back to this topic in Sect. 12.5, where some of these techniques are described
in more detail.
12.3.3 Lateral Diffusion of Lipids and Proteins
Characteristic Diffusion Coefficients
One of the compelling findings in membranes is the great variety of lateral
diffusion coefficients, which range from essentially zero to a value of about
10
−10
m
2
/s. To gain some insight into the scales on a practical level, one may
consider a molecule diffusing for a period of 10 s at a rate of 10
−11
m
2
/s.
Then the diffusion length is about 10 µm, which is roughly the diameter of
an animal cell.
12 Diffusion in Membranes 483
Diffusion in membranes is strongly dependent on phase behavior. As a
practical example one may think of diffusion in a single-component bilayer
in the vicinity of the main phase transition temperature T
M
separating the
solid-ordered phase from the liquid-disordered phase. As the temperature
is changed by a few degrees from T<T
M
to T>T
M
, the system goes
through a sudden change from a “frozen and ordered” to a liquid-like bi-
layer, and similarly the diffusion coefficient changes abruptly from 10
−20
–
10
−14
m
2
/s [23,69] to about 10
−12
–10
−10
m
2
/s [3,23, 69]. The wide range of
diffusion coefficients in the liquid-disordered phase has been obtained by a
number of different experimental techniques, and even though this may seem
surprising, the diffusion coefficient actually depends on the technique used for
the measurement. Furthermore, the diffusion coefficient depends on whether
the bilayer is free as in a liposome or supported on a solid substrate [58]
which has in some cases been found to decrease the diffusion coefficient by a
factor of 4 – 5.
This observation is essentially due to the different time and length scales
probed by different techniques. Techniques such as QENS follow the dynam-
ics of molecules in a bilayer over short time scales (less than, say, 1 ns). Thus
they examine the “short-time” behavior often denoted as “rattling in a cage”,
which corresponds to local molecular motion of a lipid within the cage of its
nearest neighbors, over a distance smaller than the size of the molecule. Rigor-
ously speaking, the motion of molecules over such short time and length scales
is not related to the lateral tracer diffusion coefficient, since the actual diffu-
sion coefficient is defined in the hydrodynamic long-time limit only. However,
one may extract a “time-dependent diffusion coefficient” by, e.g., looking at
the time-dependence of the slope of the mean square displacement. MD simu-
lation studies using this approach have led to short-time diffusion coefficients
(cf., e.g., (10.9) of Chap. 10) on the order of 10
−11
–10
−10
m
2
/s [3,40, 70].
Rather generally, the diffusion coefficients get smaller at larger time scales
measured by techniques such as FRAP. This observation is often inter-
preted as evidence of domain formation. Then one expects that |r(t)|
2
∼t
over a relatively wide time window (say, from 1 ns to 100 ns), yielding an
intermediate-time diffusion coefficient, which describes the diffusion coeffi-
cient within the domain. Only at very long times, however, when the dif-
fusion extends over domain boundaries and protein networks that tend to
slow down the diffusion coefficient, one eventually finds the hydrodynamic
diffusion behavior. The time scales that differentiate these time regimes ob-
viously depend on a system under study, but the point is that macroscopic
techniques (over time scales of microseconds and length scales of micrometers
or more) may yield diffusion coefficients that are significantly smaller than
the short-time values.
As an example of this scale-dependent diffusion behavior, Table 12.1 lists
a few recent results for the lateral diffusion of lipids in a DPPC bilayer. This is
one of the most extensively studied single-component lipid systems at present.

484 Ilpo Vattulainen and Ole G. Mouritsen
Table 12.1. Examples of lateral diffusion coefficients of lipids in single-component
DPPC (dipalmitoylphosphatidylcholine) bilayers. The time scale over which the
diffusion coefficient was determined is also given in some cases. Then “local” refers
to short times while “long range” is over a larger scale.
Technique Time scale T Diffusion coefficient Reference
MD < 10 ps 50
o
C20× 10
−11
m
2
/s [71]
MD 0.3 – 0.5 ns 60
o
C3× 10
−11
m
2
/s [3]
QENS 63
o
C40× 10
−11
m
2
/s [72]
QENS “local (max)” 60
o
C60× 10
−11
m
2
/s [64]
QENS “local (min)” 60
o
C1.5 ×10
−11
m
2
/s [64]
QENS “long range” 60
o
C1× 10
−11
m
2
/s [64]
NMR 57
o
C1× 10
−11
m
2
/s [73]
It is clear that the diffusion coefficients are reduced at long times, as the ideas
above suggest, although the effects due to domain formation expected at large
scales in many-component lipid bilayers are not included here. Further trends
of lipid diffusion have also been discussed in the literature. It has been found
that the diffusion coefficient is only weakly dependent on the chain length,
while the number of double bonds (unsaturation) has a pronounced effect on
diffusion.
Another remarkable feature associated with membranes is the role of flu-
idity for the diffusion of individual molecules. It has been found that in the
liquid-disordered phase the size of the hydrophobic part of the molecule does
not have noticeable effect on the diffusion coefficient but it is mostly deter-
mined by the properties of the head group [71, 74, 75]. The recent work by
Pampel et al. is particularly illustrating, as the pulsed field gradient NMR
studies of DOPC diffusion at 37
o
C yielded a lateral diffusion coefficient of
1.1 × 10
−11
m
2
/s. This is in full agreement with the results in Table 12.1
despite the differences in acyl chain length and saturation between DPPC
and DOPC. This finding may have major general implications. First of all,
it proposes that labeling does not influence the diffusion coefficients of the
molecules significantly, but the labeled molecules move laterally essentially
at the same rate as non-labeled ones. Recent experiments and model calcu-
lations support this view [51, 54, 74]. The same line of thought suggests that
molecules inside lipid bilayers move essentially at the same rate as the lipid
molecules in a bilayer. Again, a number of works are consistent with this
idea. They have shown that molecules close to the headgroups diffuse later-
ally approximately at the same rate as the lipids, while molecules near the
center of the membrane diffuse 3 – 4 times faster than lipid molecules [8,76,77]
(see discussion below). Further, if the hydrophobic interior of the molecule
and its size are not very important, then one might expect that the lateral
diffusion coefficient of integral proteins is roughly as large as the diffusion co-
12 Diffusion in Membranes 485
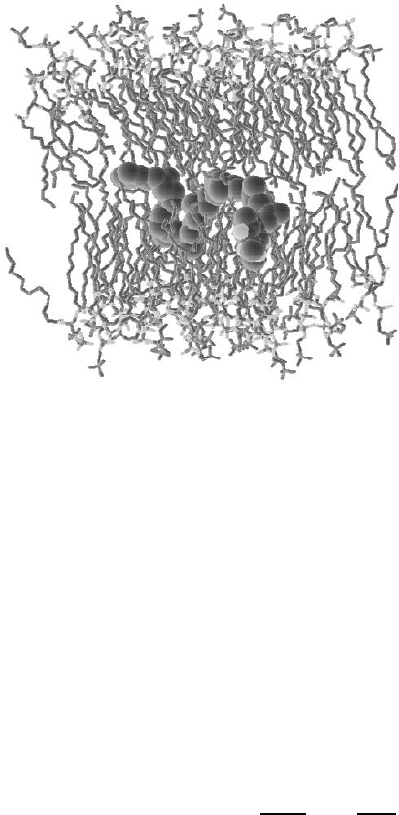
Fig. 12.5. Snapshot of an MD simulation [8] of ubiquinone (UQ-10) inside a DPPC
bilayer (water not shown). Here the ubiquinone molecule is “swimming” close to
the center of the bilayer, where the density is substantially lower compared to other
parts in a membrane. Illustration by J. Arvid S¨oderh¨all and Aatto Laaksonen.
efficient of lipid molecules. It turns out that this idea is supported by various
experiments [11].
It is indeed intriguing that the lateral diffusion coefficients of proteins
are typically from 10
−14
m
2
/s to 10
−11
m
2
/s [11, 78]. These are slower than
the diffusion coefficients of lipids, but only by a factor of ten or so. Yet
the size and mass of integral proteins are far larger than those of lipids.
Equally remarkable is the finding that the diffusion of proteins is only weakly
dependent on the physical dimensions of the protein. This is demonstrated
in the classical work by Saffman and Delbr¨uck [79], who described proteins
as cylinders with radius R, and found that their diffusion coefficient can be
described as [79]
D
=
k
B
T
4πµh
log
µh
µ
R
− γ
, (12.3)
where h is the height of the membrane in which the protein is embedded,
µ and µ
are the viscosities of the bilayer and the surrounding fluid, respec-
tively, and γ is the Euler constant. This expression is based on a number of
assumptions (including µ µ
),whicharediscussedindetailin[78].
Finally, let us briefly discuss the diffusion coefficients of molecules inside
lipid bilayers. S¨oderh¨all and Laaksonen studied the diffusion of ubiquinone
inside a DPPC bilayer in the liquid-disordered phase through MD simula-
tions [8] (see Fig. 12.5). Ubiquinone acts as a charge carrier inside lipid bi-
layers from one protein to another, and thus the diffusion properties are of
486 Ilpo Vattulainen and Ole G. Mouritsen
crucial importance in understanding the charge transfer process. S¨oderh¨all
and Laaksonen found that ubiquinone with a short tail (UQ-et) preferred a
location close to the headgroups of lipid molecules, which is probably due to
the partially hydrophilic nature of UQ-et. The diffusion of UQ-et was found
to occur with a rate being essentially the same as the diffusion of DPPC
molecules. The diffusion coefficient of ubiquinone with a long tail (UQ-10),
in turn, was found to depend on the location of the molecule in a bilayer.
Close to the membrane surface, UQ-10 and lipids diffused with similar rates,
while in the center of the membrane the diffusion of UQ-10 was found to be
almost four times faster compared to the diffusion coefficient of lipids. This
finding is consistent with earlier studies of benzene diffusion inside a DMPC
bilayer [76, 77]. Thus it seems likely that the diffusion coefficient of mole-
cules inside lipid bilayers depends on the size and density of voids, which
are greatest in the bilayer center, and that this is somehow related to the
conformational ordering of acyl chains in a membrane.
Above we have discussed the diffusion in the liquid-disordered phase. It is
worthwhile pointing out that the picture is likely to be rather different in the
solid-ordered phase, where the ordered nature of the membrane comes into
play. In the solid-ordered phase, the diffusion of molecules in the center of the
bilayer will be relatively rapid, while the diffusion close to the headgroups
will slow down due to the “frozen” nature of the bilayer.
Diffusion Mechanisms and Theoretical Frameworks
Despite the significant progress in the field, the knowledge of the microscopic
mechanisms of how molecules diffuse in membranes is scarce. It has been
proposed [65, 80] that the motion of lipids in bilayers consists of two parts:
first of local diffusion of a lipid molecule at its site, and secondly of jumps
between adjacent sites. This suggestion has been tested by a number of studies
but the overall picture is still cloudy. Essmann and Berkowitz [3] found no
evidence in MD simulations for the two regimes expected in this mechanism,
while Moore et al. found [70] that the two-step mechanism might be valid. The
MD study by Moore et al. dealt with a pure DMPC bilayer, and during a time
scale of 10 ns they found two jump events in which a lipid molecule moved
relatively rapidly over a distance of its size in the plane of the membrane. Most
lipids, however, diffused uniformly like molecules in a fluid. This suggests
that the long-range diffusion of lipid molecules in pure bilayers (in the liquid-
disordered phase) is a mixture of jumps and uniform motion.
The idea of lateral diffusion as a series of jumps from one site to another
is interesting also from a theoretical point of view, since the so-called free-
volume theory [23,78,81] is based on this idea. Originally Cohen and Turnbull
[81] derived the free-volume theory to describe diffusion of hard spheres, but
later it has been applied to a number of systems including the diffusion of
lipids in a bilayer. In this approach, one assumes that lipids spend long periods
of time at their sites, and long-range diffusion takes place only if there is a