Хаитов P.M., Игнатьева Г.Л. Иммунология
Подождите немного. Документ загружается.

-2
α1
y<
β1
β2
MHC-I
MHC-II
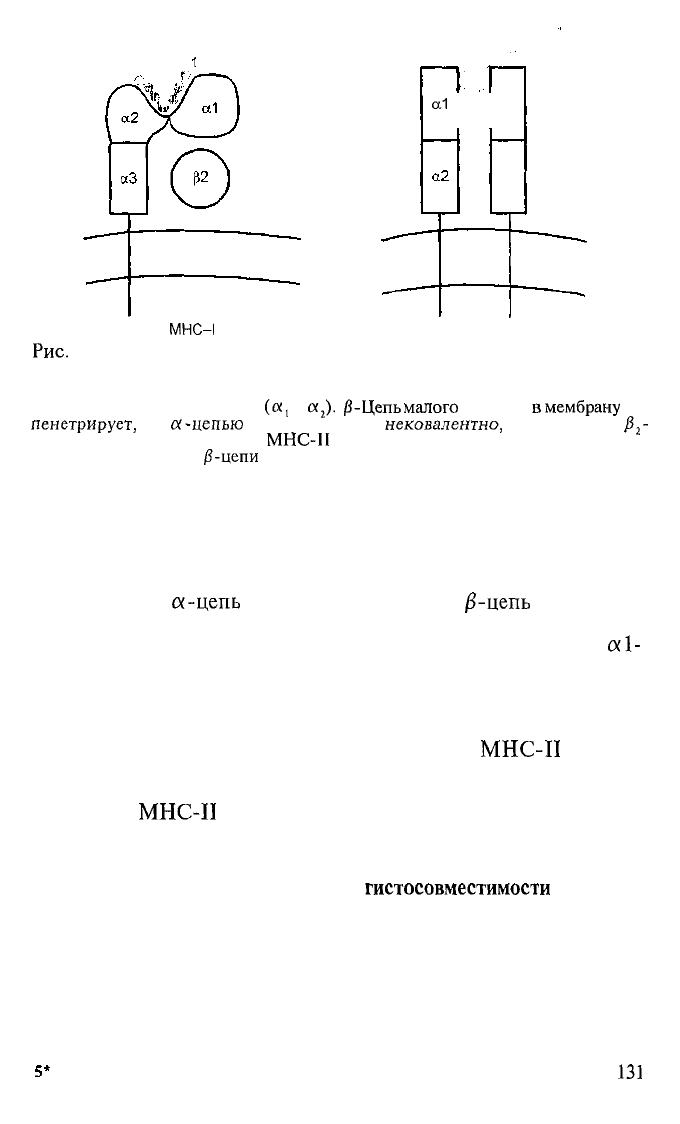
Рис.
5.7. Структура молекул MHC-I и II (схема).
Молекулы MHC-I — трансмембранные гетеродимеры: α-цепь большая, имеет
три внеклеточных домена. «Клефт» для связывания пептида формируют два
терминальных домена «-цепи
(ct
t
и
сх
2
).
β-Цепь
малого
размера,
в
мембрану
не
пенетрирует,
с
α-цепью
ассоциирована
нековалентно,
ее называют
β
2
-
микроглобулином. Молекулы
МНС-П
также трансмембранные гетеродимеры.
Полипептидные а- и
/?-цепи
равновелики, обе пенетрируют мембрану, у обе-
их по два внеклеточных домена и «клефт» для связывания пептидов формируют
внешние домены обеих цепей. 1 — пептид-антиген в комплексе с молекулами
MHC-I; 2 — пептид-антиген в комплексе с молекулами МНС-П.
ные молекулы — гетеродимеры из двух трансмембранных гли-
копротеинов:
α-цепь
(мол. масса 34 000) и
/3-цепь
(мол. мас-
са 29 000). Внеклеточная часть каждой цепи имеет два домена.
Пептидсвязывающий клефт (cleft) формируют совместно
otl-
и β\-домены и в отличие от «клефта» MHC-I «клефт» MHC-
II открыт с обеих сторон (рис. 5.7). Стереохимическая откры-
тость «клефта» МНС-П позволяет связывать более длинные
пептиды, чем в случае MHC-I, — до 30 остатков аминокис-
лот. Однако чаще всего из комплексов с
МНС-И
удается
диссоциировать пептиды размером 13—17 остатков аминокис-
лот. Якорные остатки для большинства изученных аллельных
вариантов
МНС-И
находятся в позициях 1; 4; 6 и 9.
5.7. Механизмы образования комплексов пептидов-антигенов
с молекулами главного комплекса
гистосовместимости
В любой эукариотической клетке есть две «зоны», или, как
их принято называть от английского, компатменты. Одна зона
связана с внешней внеклеточной средой и это зона мембран-
ных структур и везикул, включающая в себя аппарат Гольд-
жи, эндоплазматический ретикулум, лизосомы, эндосомы и
фагосомы. Пептиды, образующиеся в данной зоне, обслужи-
131

воют
синтезируемые клеткой молекулы МНС-П. Происхожде-
ние пептидов везикул внеклеточное: они образуются в резуль-
тате протеолиза белков, захваченных клеткой посредством эн-
доцитоза или фагоцитоза. Эндосомы, или фаголизосомы, сли-
ваются с мембранными структурами, на которых фиксируют-
ся вновь синтезированные на полирибосомах молекулы а- и
β -цепей МНС-П и еще одна так называемая инвариантная
полипептидная цепь — П. Эта
Ii-цепь
«прикрывает» МНС-П
от связывания с пептидами внутри эндоплазматического ре-
тикулума и в дальнейшем обеспечивает экспозицию молекул
МНС-И внутрь везикул — эндосом, или фаголизосом. Комп-
лекс из 3 тримеров
αβΙΊ
в эндоплазматическом ретикулуме
удерживается связью с кальнексином. В мембранных внутри-
клеточных структурах есть специальный компартмент М-Н-С
(МНС class II compartment), в который физически упираются
эндосомы и лизосомы с поглощенным внеклеточным содер-
жимым. И только с участием пептида молекула МНС-Н при-
нимает правильную
конформацию,
продвигается к мембране и
экспрессируется на ней. «Чистые» молекулы МНС I и II класса
на мембране клетки не экспрессируются. Если клетка не рас-
полагает достаточным количеством чужого пептидного мате-
риала, то молекулы МНС-П связывают короткий фрагмент,
образующийся при протеолитическом расщеплении инвариант-
ной цепи — CLIP (class II linked
invariant-chain
peptide).
В наслоении пептида на МНС-П участвует еще одна молеку-
ла — HLA-DM. HLA-DM на какое-то время стабилизирует пу-
стые молекулы МНС-И (без пептида). Отсюда понятно, по-
чему представление антигена именно молекулами
МНС-П
осу-
ществляется при развитии защитных иммунных реакций на
внеклеточные и везикулярные инфекции. Поскольку комплексы
антигенов-пептидов с молекулами
МНС-Н
распознают исклю-
чительно
CD4
+
Т-лимфоциты, то и в защите от внеклеточных
и везикулярных инфекций главную роль играют реакции, обеспе-
чиваемые именно CD4 Т-лимфоцитами (рис. 5.8).
Вторая «зона» в клетке — зона цитозоля — неструктури-
рованного мембранами внутриклеточного содержимого. Цито-
золь непосредственно сообщается через крупные ядерные
поры с содержимым ядра. В ядре идут процессы транскрип-
ции РНК с ДНК (как нормальные клеточные, так и вирус-
ные при вирусных инфекциях клеток). РНК из ядра транспор-
тируется для трансляции белка в цитозоль на полирибосомы,
и для фолдинга (принятия правильной конформации) белков
под контролем шаперонов («катализаторов» принятия поли-
пептидной цепью биологически значимой конформации)
полипептидные цепи направляются в специальные участки
эндоплазматического ретикулума. Лишние, испорченные, не-
правильно конформированные белки постоянно расщепляют-
ся в цитозоле так называемыми протеасомами.
Протеасомы
—
132

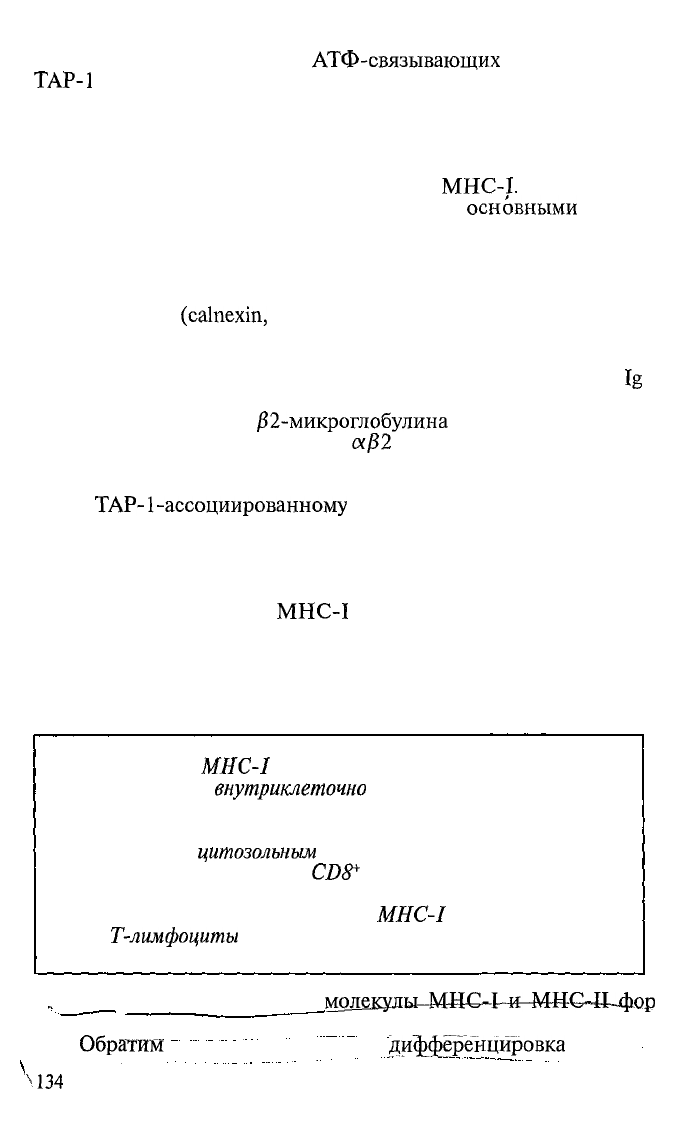
Рис. 5.8. Строение клетки и «зоны обслуживания» молекулами MHC-I
и II (схема).
I
— зона (или
компартмент)
мембранных структур клетки (везикулы, эндо-
плазматический
ретикулум, аппарат Гольджи), эта зона непосредственно со-
общается с внеклеточной средой; 2 — зона (или компартмент) цитозоля
непосредственно сообщается с внутриядерным содержимым; 3 — протеасомы
в
питозоле;
4 — фагосомы; 5 — фаголизосомы сливаются с везикулами,
содержащими несконформированные молекулы МНС-И; 6 — комплекс МНС-
II — пептид «внеклеточного» происхождения экспрессирован на мембране клет-
ки; 7 — несконформированные молекулы MHC-I в зоне досягаемости пепти-
дов — продуктов протеасом; 8 — комплекс MHC-I — пептид «внутриклеточного»
происхождения экспрессирован на мембране клетки.
это
мультипротеазные
комплексы из 28 субъединиц, каждая
из которых имеет молекулярную массу 20 000 или 30 000. Два
из 3 вариантов субъединиц протеасом — LMP2 и LMP7 —
кодируются генами, расположенными внутри комплекса
мне.
Протеасомы осуществляют зависимую от убиквитина
(ubiquitin) деградацию белков цитозоля. И именно пептиды,
образующиеся в результате работы протеасом, поступают в
«поджидающие» их в нужном месте эндоплазматического ре-
тикулума еще несконформированные молекулы
MHC-I:
всту-
пая в связь с пептидом, молекулы
MHC-I
принимают био-
логически правильную конформацию и направляются для
экспрессии на клеточную мембрану. Исследование мутаций и
генетических дефектов, приводящих к иммунодефицитным
133
состояниям, выявило зависимость представления антигена
молекулами MHC-I от двух
АТФ-связывающих
полипептидов
ТАР-1
и ТАР-2 (Transporters associated with antigen processing).
В мембране эндоплазматического ретикулума эти два полипеп-
тида формируют гетеродимер, ориентированный гидрофобны-
ми участками в полость ретикулума, а АТФ-связывающими
доменами — в сторону цитозоля. Именно они доставляют
пептиды для связывания с молекулами
MHC-I.
ТАР имеют
сродство к пептидам с гидрофобными или
основными
остат-
ками аминокислот на С-конце. Гены ТАР-1 и ТАР-2 распо-
ложены внутри комплекса МНС.
Вновь синтезированная «-цепь MHC-I вступает в связь с
мембранным протеином эндоплазматического ретикулума
кальнексином
(calnexin,
мол. масса 88 000), который обеспе-
чивает частичный фолдинг «-цепи. Кстати, тот же кальнек-
син образует комплексы с вновь синтезированными и частич-
но сконформированными полипептидами МНС-П, TCR,
Ig
(т.е. всеми иммунологически значимыми пептидами). При при-
соединении к «-цепи
/?2-микроглобулина
связь с кальнекси-
ном диссоциирует, и комплекс
αβ2
присоединяется для даль-
нейшего прохождения процесса приобретения конформации
к кальретикулину (calreticulin) эндоплазматического ретикулу-
ма и
ТАР-1-ассоциированному
протеину тапасину (tapasin).
Таким образом создаются условия для соединения пептида-
антигена с принимающим конформацию комплексом MHC-I.
При генетических дефектах в ТАР молекулы МНС деградиру-
ют внутри клетки, не попадая на клеточную мембрану. В нор-
ме в клетке молекулы
MHC-I
присутствуют в избытке по
отношению к пептидам. Поэтому в клетке при вирусных ин-
фекциях довольно быстро пептиды вирусного происхождения
оказываются представленными на поверхности зараженных
клеток в составе комплексов с MHC-I.
Молекулы
MHC-I
представляют для распознавания
Т-лимфоцитам
внутриклеточно
синтезированные пептид-
ные антигены. В плане защиты от инфекций этот меха-
низм работает применительно в первую очередь к вирус-
ным, а также
цитозолъным
бактериальным внутриклеточ-
ным инфекциям. Поэтому
CD8*
Т-лимфоциты «рассчита-
ны» в первую очередь на обеспечение противовирусной за-
щиты, так как в комплексе с
MHC-I
именно и только
CD8
Т-лимфоциты
распознают пептидные антигены.
. В отсутствие инфекций
мшщкулы-фр
) мируют комплексы с эндогенными (собственными) пептида-
ми.
Обратим
внимание на то, что
диф^фёренцйровка
Т-лим-
434
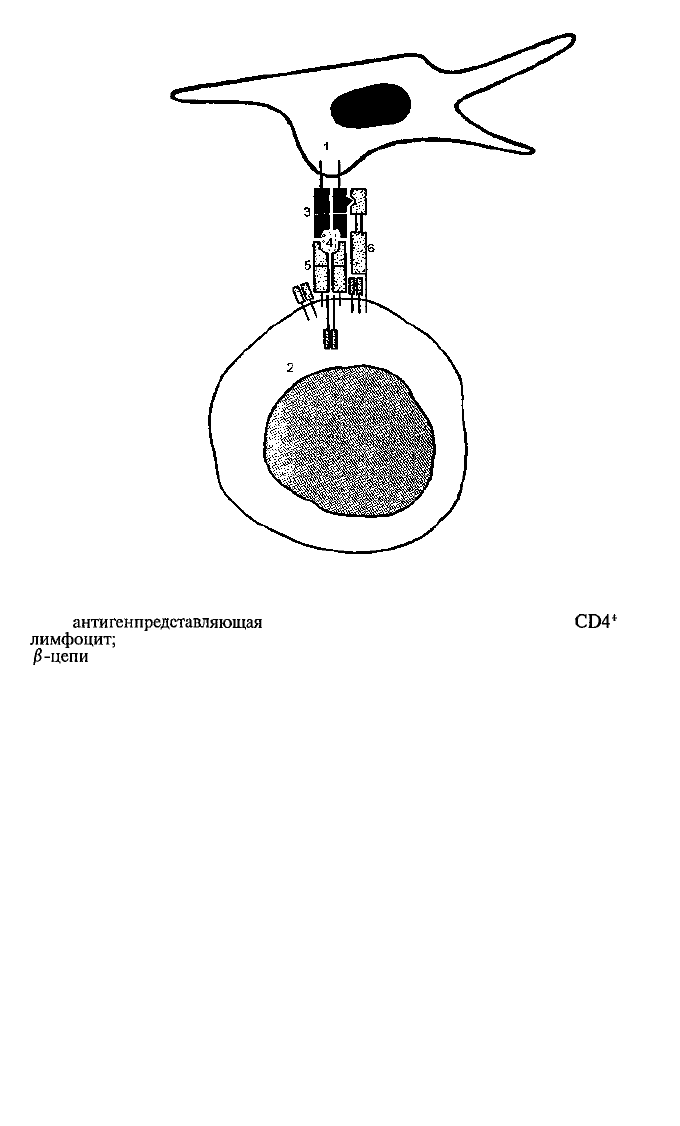
Рис. 5.9. Взаимодействие TCR с комплексом молекулы МНС-пептид на
антигенпредставляющей клетке.
1 —
антигенпредставляющая
клетка (например, дендритная); 2 —
CD4*
Т-
лимфоцит;
3 — молекулы МНС-И; 4 — пептид (собственно антиген); 5 — а- и
β-цепи
TCR; 6 — молекула CD4.
фоцитов в тимусе в норме в стерильном изнутри организме
идет при взаимодействии отбираемых TCR со своими пепти-
дами, а не на отборе «своего и чужого». Поэтому естествен-
но, что среди случайно формируемого разнообразия вариан-
тов антигенсвязывающих структур даже после позитивной и
негативной селекции тимоцитов среди выходящих на перифе-
рию Т-лимфоцитов обязательно найдутся TCR, способные спе-
цифично связываться, наверное, со всеми собственными «ан-
тигенами». Опыт показывает, что так оно и есть. Но деструк-
тивный иммунный ответ на собственные антигены в норме не
развивается, и в дальнейшем мы обсудим то немногое, что
известно о механизмах периферической толерантности.
Стабильное связывание антигена молекулами МНС внутри
клетки при формировании собственной конформации, а не
простая сорбция из раствора антигена снаружи уже экспрес-
сированными на мембране молекулами МНС, обеспечивает
направление Т-лимфоцитарного иммунитета на реально ин-
135
фицированные клетки тела. Иммунное воспаление, управляе-
мое активными Т-лимфоцитами, — это наиболее «энергич-
ное» воспаление по сравнению с другими вариантами и та-
кое воспаление нельзя «натравливать» на ложные мишени,
иначе организм будет не направленно избавляться от инфек-
ции, а страдать от неоправданно разлитой альтерации тканей.
Взаимодействие TCR с комплексом пептид — MHCI/II
изображено на рис. 5.9.
5.8. Суперантигены
В борьбе за свое существование инфекционные микроорганиз-
мы «научились» синтезировать некие вещества, которые по-
ступают с «нашими» Т-лимфоцитами «не по правилам». Эти
вещества называют суперантигенами. Когда они попадают в
организм, то последний не может встретить их попыткой
распознавания специфическим TCR потому, что суперанти-
гены благодаря своим особым химическим свойствам способ-
ны присоединяться
стршси_юг_етки_£1.е.
без процессинга в
антигенпредставляющих
клетках)~с
одной стороны к молеку-
лам МНС-И, но не в области
«клефта
для пептида», а «сбо-
ку»^
как правило, к «-цепи, и одновременно к V-области
•"ТТ^цепи
TCR на Т-лимфоците.
Это связывание вызывает активацию Т-лимфоцитов, но не
антигенспецифичного клона, а поликлональную. Обычно су-
перантигены активируют 2—20 % периферических
CD4
+
T-
лимфоцитов. В результате многие Т-лимфоциты продуцируют
и секретируют активационные цитокины, что вызывает син-
дром общей интоксикации в организме. А затем активирован-
ные Т-лимфоциты погибают путем апоптоза. Таков закон лим-
фоцитарного иммунитета: на свой ли антиген или неспеци-
фически, но единожды активированный лимфоцит обречен на
апоптоз (феномен AICD
—
activation-induced cell
deatrT'"—
индуцированной активацией клеточной смерти). (Феномен
иммунологической памяти — особое явление, и хотя оно
исключительно характерно именно для
лимфоцитарного
им-
мунитета, механизмы его непонятны до настоящего времени.)
Но фактом является то, что после поликлональной актива-
ции лимфоцитов наступает физический дефицит лимфоцитов,
следовательно, иммунодефицит.
Известны суперантигены бактериального и вирусного про-
исхождения, такие как энтеротоксины стафилококков (SE),
токсин синдрома токсического шока (TSST-1 — toxic shock
syndrome
toxin-1),
мембранный протеин вируса опухолей мо-
лочных желез
мышей,
jiaii
_^
вируса"
бешенства, вирусов
иммунодефицита
человека~{КИЧ)
и животных и многие дру-
гие. Клиника пищевых отравлений, например, обязана глав-
136
ным образом действию SEs на Т-лимфоциты. Иммунодефицит
в ранние сроки (и в поздние тоже) ВИЧ-инфекции в значи-
тельной мере объясняется лимфотоксическим действием су-
перантигенов вируса ВИЧ.
5.9. Генетический полиморфизм главного комплекса
гистосовместимости
Итак, в случае Т-лимфоцитов пептид-антиген связывается не
только с рецептором Т-лимфоцита для антигена, но и (при-
чем по времени «до» Т-лимфоцита) с молекулами МНС ан-
тигенпредставляющих клеток. Разнообразие TCR природа со-
здает комбинаторикой сегментов ДНК — уникальным
процес-
сом рекомбинации ДНК в соматических клетках. А как моле-
кулы МНС приспособлены к открытому множеству заранее
неизвестных антигенов? Иначе, чем
TCR
и иммуноглобули-
ны.
Рекомбинации.
ДНК в
генах]УШС_дет\_Каждый
человек
наследует.о"пределенный~набоТГТенов
МНС,
один и
тот
же,
на всю
жизнь^_Гены
УЩ.С^-кодашшамшиы^
Это значит,
что
на
равных экспрессируются гены обеих гомологичных хромо-
сом — от матери и от отца.
Гены МНС полигенны, т.е. существует
-не
один, а несколь-
ко локусов как I, так и II класса внутри комплекса
МНС.
Еще есть и так называемые неклассические гены, правильнее
их было бы назвать не принадлежащими к ранее охарактери-
зованным I и II классам МНС. Сколько же генов МНС? Этот
участок генома картирован не полностью, но то, что извест-
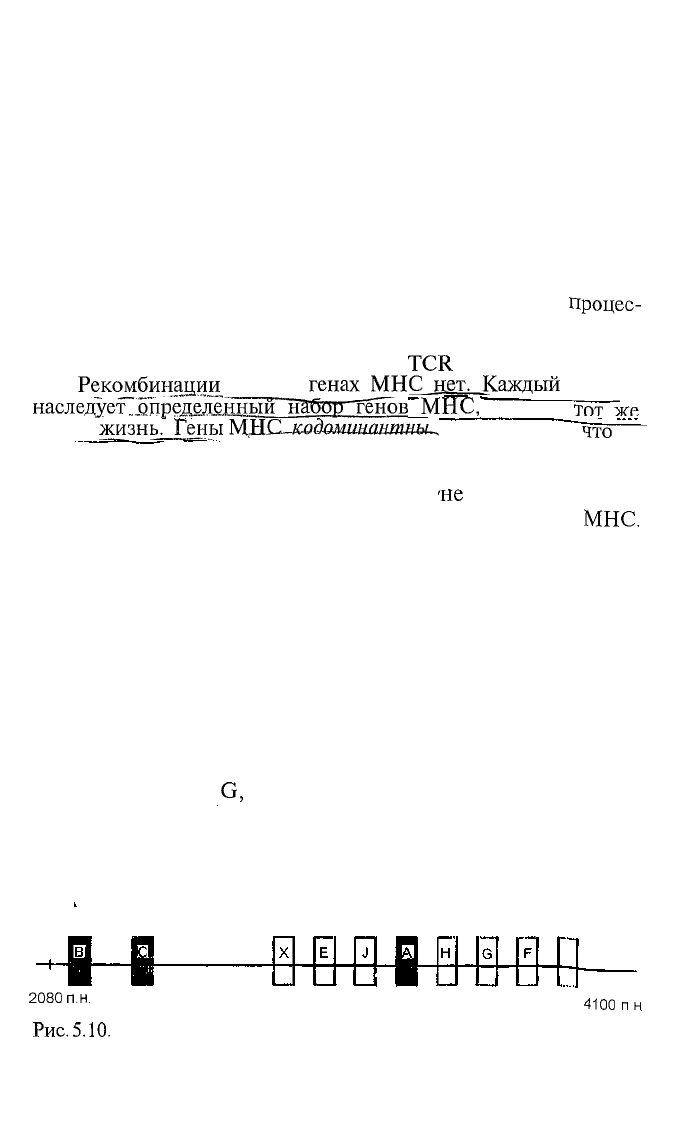
но о нем, показано на рис. 5.5 и 5.10.
Таким образом, генов MHC-I по 3 на каждой из гомоло-
гичных хромосом (А, В, С), следовательно, в сумме их 6. Ге-
нов МНС-П также по 3 на каждой из гомологичных хромосом
(DP, DQ, DR), следовательно, в сумме их тоже 6. Если у
матери и отца нет одинаковых аллелей, то каждый человек
имеет 12 генов МНС I и II класса, вместе взятых. Некласси-
ческих генов МНС в области расположения генов I класса
описано 6 (Е, F,
G,
Н, J, X). Возможно, будут открыты и
новые гены. Неклассические гены МНС, видимо, неполиморф-
ны или не столь полиморфны, как гены I и II классов.
Известны гены за пределами МНС, кодирующие молеку-
лы, которые обеспечивают представление антигенов Т-лим-
2080 пн
4100
пн
Рис.
5.10. Расположение неполиморфных аллелей области МНС челове-
ка (схема).
137
фоцитам. Примером такой молекулы (и гена соответственно)
является
CD1,
способная связывать и представлять Т-лимфо-
цитам небелковые антигены (например, миколевую кислоту
и липоарабиноманнан оболочки микобактерий).
Описаны неполиморфные молекулы и из области МНС-
II — DM и DQ. Функции HLA-DM в какой-то мере извест-
ны, и о них мы уже упоминали (стабилизация молекул МНС-
II внутри клетки во время «ожидания» смыслового пептида и
«катализ» присоединения к МНС-П смыслового пептида, а
также генерация «протезного»
CLIP-пептида).
Гены полипеп-
тидных цепей, формирующих молекулу DQ, экспрессируют-
ся, по-видимому, только в тимусе и В-лимфоцитах, но фун-
кции этой молекулы неизвестны.
Полиморфизм классических генов МНС (I и II классов)
означает наличие в популяции множества аллелей — ва-
риантов одноименного гена у разных особей.
Поэтому разные люди, неродственники, имеют различные
аллели одноименных генов и, следовательно, разные белки
МНС на мембранах своих клеток. Вероятность существования
одинаковых аллелей у неродственников, тем более совпаде-
ния по всем или многим локусам тем меньше, чем больше
полиморфизм в популяции. Полиморфизм генов МНС признан
наибольшим из всех известных генов человека.
Полиморфизм этих генов в популяции выгоден для выжи-
вания вида: не у одного, так у другого человека в популяции
организм может быть сумеет представить Т-лимфоцитам сво-
им вариантом молекул МНС антигены того или иного (в том
числе и из вновь возникающих) инфекционного микроорга-
низма. Но вот каждый конкретный человек либо сумеет, либо
нет воспринять конкретный микробный антиген и если не
сумеет, то не сможет защититься от данного микроба иммун-
ным ответом и инфекция может убить его.
Конкретные варианты МНС закрепляются в эволюции
естественным отбором (в отличие от TCR и
Ig)
и каж-
дая отдельная особь оказывается приспособленной к реги-
онарным видам и штаммам инфекционных микроорганиз-
мов, на защиту от которых шел отбор МНС у предков.
Для отдельных локусов МНС известно более сотни аллель-
ных вариантов, и поскольку далеко не каждый человек на
Земле типирован по МНС, привести точное число аллелей
138
нельзя. Международные банки данных располагают сведения-
ми о не менее чем 60 аллелях HLA-A, 111 аллелях HLA-B,
37 аллелях HLA-C, 62 аллелях
HLA-DP/5,
8 аллелях HLA-
DPa,
25 аллелях
HLA-DQjS,
16 аллелях
HLA-DQa,
122 ал-
лелях
HLA-DR^,
1 аллеле
HLA-DRa.
Каждый отдельный аллельный вариант молекулы МНС
преимущественно связывает пептиды с определенными якор-
ными аминокислотными остатками.
v
От 1 до 10 % Т-лимфоцитов любого здорового организма
реагируют активацией при контакте с аллогенными по МНС
клетками. На любой случайный антиген, однако, реагирует
гораздо меньшее число Т-лимфоцитов антигенспецифичных
клонов. Столь высокий уровень аллореактивности Т-лимфоци-
тов оставался непонятным (учитывая отсутствие в природе
пересадок органов и тканей) до открытия двойного распоз-
навания Р.Цинкернагелем и П.Дохерти. Тогда стало возмож-
ным предполагать, что аллореактивность — результат перекре-
стной реактивности с не своими МНС TCR, отобранных при
позитивной селекции в тимусе на связывание со своими МНС.
Поэтому приходится признать, что отторжение аллогенного
трансплантата происходит в результате того, что 1—10 %
Т-лимфоцитов
реципиента «ошибаются», принимая донорские
МНС за
свой(гТё"ре^рё1:тно~^^хируют),
тогда как
Э9—90Г%~
Т-лимфоцитов
реципиента
«НЕПЗЙДЯТ»
чужой
трансплантат,
поскольку на нем не те МНС, на связывание с которыми
прошла позитивная селекция тимоцитов. ^
Почему же число генов МНС I + II у индивидуума 12 и не
больше? Ведь чем больше было бы генов МНС, тем большее
разнообразие антигенов могли бы представлять антигенпред-
ставляющие клетки для распознавания Т-лимфоцитам. Огра-
ничение числа генов МНС можно объяснить механизмами по-
зитивной и негативной селекции тимоцитов на связывание с
МНС, представленными в тимусе во время и на месте лим-
фопоэза Т-лимфоцитов.
Исследования показывают, что из каждого подмножества
тимоцитов, оставленных живыми при позитивной селекции на
связывание с конкретным вариантом молекулы МНС, 5 %
будет элиминировано при негативной селекции как слишком
аутореактивные, т.е. каждый вариант молекул МНС «стоит
жизни» 5 % тимоцитов. На 12 вариантов молекул МНС (если
считать только I и II классы) в тимусе должно быть ликви-
дировано 12 х 5 = 60 % тимоцитов. Реально в тимусе погиба-
ет более 95 % тимоцитов и лишь менее 5 % выходят в пери-
ферические лимфоидные органы. Видимо, в эволюции увели-
чивать полигенность МНС больше было нельзя.
Есть еще такой термин, как
_«гены
MHCzIlI».
Это область
генома внутри комплекса МНС между I и И, в которой кар-
тированы гены, кодирующие синтез следующих белков:
139
1) компонентов системы комплемента С4А и С4В, С2,
фактора В;
2) цитокинов — фактора некроза опухолей
(TNF-oi)
и
лимфотоксина (LT);
3) гены
21-гидроксилазы
— CYP 21А, CYP 21B (фермен-
та, участвующего в биосинтезе стероидных гормонов).
До настоящего времени неизвестно, что кодирует (и ко-
дирует ли) значительная часть ДНК этой области.
5.10.
Антигенпредставляющие
молекулы «не МНС» — CD1
В организме у человека есть семейство, состоящее из 5 близ-
кородственных генов, которые неполиморфны и кодируют со-
ответственно 5 изоформ мембранных белков, по структуре
похожих на MHC-I. Это кластер CD1:
CDla,
CDlb,
CDlc,
CD
Id,
CDle;
a, b и с обладают значительной гомологией и
составляют одно подсемейство, d и е — второе. Молекулы
CD1
—
гетеродимеры,
состоящие из
α-цепи
с молекулярной
массой 45 000 и
)3
2
-микроглобулина.
Антигенсвязывающая
«впадина» на
α-цепи
уже и глубже, чем у молекул MHC-I,
но главное, что место связывания антигена имеет большое
химическое сродство к гидрофобным лигандам. Недавно по-
казано, что антигены, которые связывают молекулы кластера
CDla, b и с, имеют гидрофобную область (цепи жирных кис-
лот) и гидрофильную область из полярных или заряженных
групп (гетерополисахариды). Например,
CD1
связывают липог-
ликаны микобактерий, состоящие из гидрофобных липидсо-
держащих фосфатидилинозитольных групп, связанных с круп-
Таблица 5.5. Экспрессия различных изоформ молекулы CD1 на раз-
ных клетках
Изоформа
Человек
1. CDla
2. CDlb
3. CDlc
4.
CDld
5. CDle
Мышь
6. CDl.l
7.
CD
1.2
На каких клетках
экспрессирована
Дендритные клетки; клетки Лангерганса;
кортикальные тимоциты
Дендритные клетки; кортикальные тимо-
циты
Дендритные клетки; клетки Лангерганса;
кортикальные тимоциты;
В-лимфоциты
В-лимфоциты; эпителиальные клетки
кишки; другие
нелимфоидные
клетки (?)
Неизвестно
Тимоциты; В-лимфоциты; Т-лимфоциты;
макрофаги; дендритные клетки; энтероци-
ты; клетки печени
Неизвестно
140