Хаитов P.M., Игнатьева Г.Л. Иммунология
Подождите немного. Документ загружается.

отличающиеся варианты иммуноглобулинов в пределах одноимен-
ного изотипа — это аллельные варианты, или аллотипы, имму-
ноглобулинов. Факт существования
аллотипов
свидетельствует о
некотором генетическом полиморфизме внутри вида по ло-
кусам С-генов как легких, так и тяжелых цепей.
\ Антисыворотки против уникальных вариабельных участков
1
молекул иммуноглобулинов называют антиидиотипическими, а
Соответствующие
эпитопы в молекуле антител — идиотопом
антитела
—
(idious
— уникальный, не такой, как другие).
Таким образом, идиотип антитела — это вариант уникального
штигенсвязывающего участка молекулы иммуноглобулина.
Перечислим свойства человеческих иммуноглобулинов раз-
ых классов (табл. 4.2).
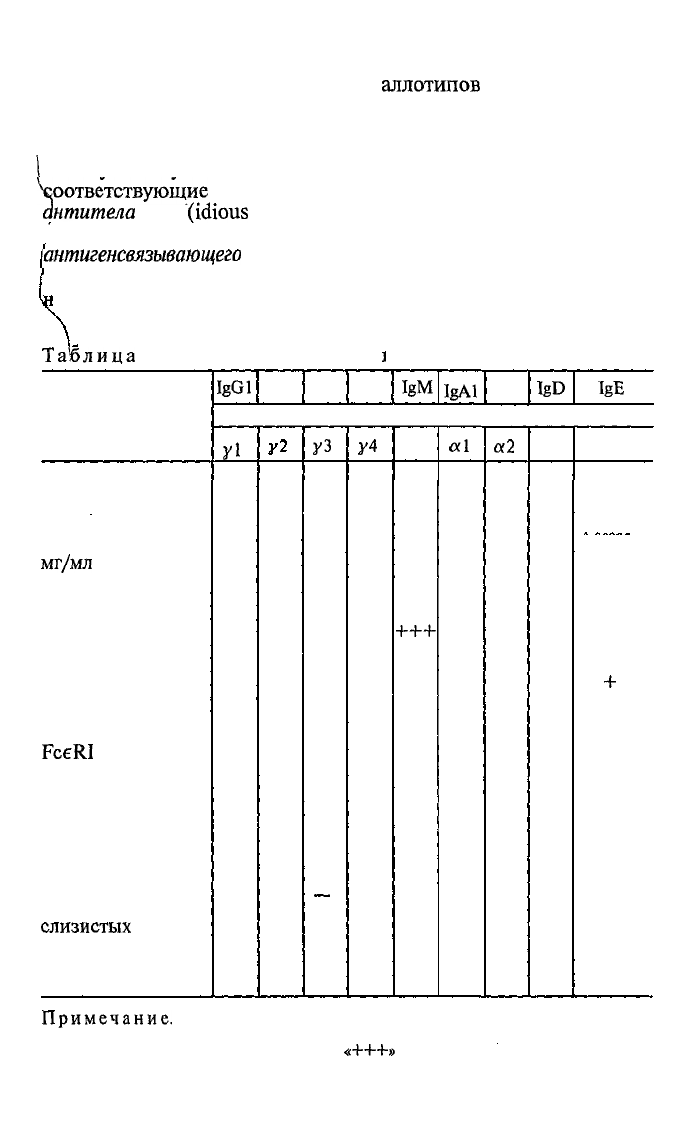
Таолица
4.2. Свойства человеческих
ι
Свойство
Молекулярная
масса, х 1000
Концентрация в
сыворотке крови,
мг/мл
Время полураспада
в крови, сут
Активация комп-
лемента по клас-
сическому пути
Связывание с
Fc-рецептором
фагоцитов
Связывание с
FceRI
тучных кле-
ток и базофилов
Способность
нейтрализовать
инфекционность
вирусов и
бактерий
Экзосекреция
через эпителий
слизистых
оболочек
Проникновение
через плаценту
IgGl
lgG2
п
146
9
21
++
+++
—
++
—
+++
146
3
20
+
—
—
++
—
+++
IgG3
IgG4
иммуноглобулинов
IgM
Тяжелая
уз
165
1
7
+++
++
—
++
__
+++
У4
146
0,5
21
—
—
—
++
_
+++
μ
970
1,5
10
+++
+
—
—
+
+
—
IgAl
lgA2
IgD
IgE
цепь
αϊ
160
3
6
—
—
—
++
+++
—
α2
160
0,5
6
—
—
—
++
+++
—
δ
184
0,03
3
—
—
—
—
—
—
e
188
0,00003-
0,00005
2
—
+
+++
—
++
—
Примечание.
Количественная выраженность того или иного свойства в
данном случае представлена в условных знаках: «—» — отсутствие; «+» —
немного; «++» — больше, чем «+»;
«+++»
— больше, чем «++».
91

Низкие концентрации, например,
IgA
в крови не означа-
ют, что организм продуцирует IgA меньше, чем
IgG.
Скорее
наоборот: суточная продукция IgA возможно максимальна
среди прочих изотипов и составляет около 3 г, но его физи-
ологическое место не в крови, он секретируется из внутрен-
ней среды во внешнюю — в слизистые экзосекреты и таким
образом является фактором специфической иммунной защи-
ты внутренней среды, вынесенным во внешнюю за пределы
покровных тканей. Кстати, в работах последних лет появились
данные, свидетельствующие о том, что и иммуноглобулины
другого изотипа —
IgE
более чем на 90 % секретируются в
слизистый экзосекрет ЖКТ. (Подробнее о функциональных
свойствах антител разных классов рассказано в главе 8.)
4.6. Дифференцировка В-лимфоцитов
Дифференцировка В-лимфоцитов из общей лимфоидной клет-
ки-предшественницы — потомка стволовой кроветворной
клетки состоит из следующих этапов и процессов:
• развитие молекулярно-генетического аппарата, обеспе-
чивающего биосинтез иммуноглобулинов — это пере-
стройка генов иммуноглобулинов (обеспечивающая раз-
нообразие антигенсвязывающих областей иммуноглобу-
линов) и настройка этих генов на продуктивную интег-
рацию в клеточный метаболизм;
• экспрессия генов молекул, обеспечивающих проведение
сигнала с иммуноглобулинового рецептора для антиге-
на внутрь клетки;
• экспрессия генов мембранных молекул, необходимых для
участия В-лимфоцита во взаимодействиях с другими клет-
ками, в первую очередь с Т-лимфоцитами и фоллику-
лярными дендритными клетками. Это молекулы CD40,
МНС-И, CD45, рецепторы для
цитокинов-факторов
роста
(IL-7 во время лимфрпоэза, IL-2 во время иммуногенеза);
• для эффективного функционирования В-лимфоцитов "су-
щественна экспрессия на мембране корецепторов
CD
19,
CD20 и CD21. Не случайно именно эти мембранные
молекулы используются как маркеры для определения
содержания В-лимфоцитов лабораторными методами
идентификации клеток.
Прежде чем описать последовательность событий диффе-
ренцировки В-лимфоцитов, скажем о существовании двух
известных на настоящее время субпопуляций В-лимфоцитов —
В-1
и В-2.
В_-2;ЛИмфрциты
— это те лимфоциты, про кото-
рые знали
раньше.
В-1-лимфоциты
стали известны относи-
тельно недавно и «проявили» они себя при детальных анали-
зах определенных клинических случаев лейкозов.
В-1-лимфо-
92

циты несут мембранный маркер, которого нет на
В-2-лим-
фоцитах, — это
молекула_СЩ.
Та же молекула экспрессиру-
ется и на
некоторой
части
ТЧлимфоцитов.
В-1-лимфоциты
поддерживают свою физиологическую ре-
генерацию в течение всей жизни из отдельной клетки-пред-
шественницы, пул
кото£ОЙ
jM^pocjmix
переполняется
за счет
общей
стволовой
кровётворнои""клетки
костного'
мозгаГ
Эта
отдельйая
клетка-предшественница
отселяется из кроветвор-
ной ткани на свою анатомическую территорию — в
брющ-
ную и плевральную полости — еще в эмбриональном перио-
де.
Йтак7
место"
обитания
В-1-лимфоцитов
—
прибарьерные
полости. В-1-лимфоциты значительно отличаются от В-2-лим-
фоцитов по
антигенраспознавательньш
способностям проду-
цируемых антител. Антитела, синтезированные
В-1-лимфоци-
тами, не имеют значительного разнообразия вариабельных
участков молекул иммуноглобулинов, но, напротив, ограни-
чены в репертуаре распознаваемых антигенов, и эти антиге-
ны — наиболее распространенные соединения клеточных сте-
нок бактерий. Все В-1-лимфоциты представляют собой как бы
один не слишком
специализированный,
но определенно
ори-
ентированный
(антибдкхержадьный)
клон.
Антитела^
продуци-
руемые
В-1-лимфоцитами,
почти исключительно
IgM;'.
пере-
ключение классов
иммуноглобулинов
"в
"В-1-лимфоцитах
не
«предусмотрено»!
1
Таким образом, В-1-лимфоциты — «отряд»
противобактериальных «пограничников» в прибарьерных по-
лостях, предназначенных для быстрой реакции на «просачи-
вающиеся» через барьеры инфекционные микроорганизмы из
числа широко распространенных. В сыворотке крови здорового
человека
большая
часть иммуноглобулинов — продукт синте-
за как раз
В-1-лимфоцитами,
т.е. это относительно полиспе-
цифичные иммуноглобулины антибактериального назначения.
В-2-лимфоциты
— это лимфоциты, характеризующиеся
широким разнообразием антигенраспознающих участков мо-
лекул продуцируемых ими иммуноглобулинов. Они проходят
свой лимфопоэз в раннем эмбриогенезе на территории пече-
ни, затем исключительно на территории костного мозга, а
свой иммуногенез — строго в фолликулах периферических J
лимфоидных органов. В лимфопоэзе В-2-лимфоцитов выделяют/
6 этапов: [общая лимфоидная клетка-предшественник] -*
ранняя
npo-JB-клетка
-+
поздняя про-В-клетка -* большая
пре-В-клетка
->•
малая пре-В-клетка
—>·
незрелая В-клетка -*
зрелая неиммунная В-клетка (выходит из костного мозга в
периферическую лимфоидную ткань).
Клетки стромы костного мозга обеспечивают оседлость
развивающихся
В-лимфоцитов
за счет взаимодействия опре-
деленных молекул межклеточной адгезии и факторы роста для
необходимого числа циклов пролиферации. Как и во всех слу-
чаях клеточной дифференцировки, самые ранние механизмы
93

комитации к данному пути, а не к другому, неизвестны. Но
ряд маркеров движения по пути В-лимфопоэза известны.
На ранней лимфоидной клетке-предшественнице экспрес-
сируются несколько молекул адгезии, обеспечивающих осед-
лость в течение необходимого периода времени в костном
мозге, среди них VLA-4 (very late antigen-4 — очень поздний
антиген 4), лигандом которого на клетках стромы является
VCAM-1
(vascular cell adhesion
molecule-1
— молекула адге-
зии 1 к стенке сосуда). На ранней про-В-клетке, кроме мо-
лекул адгезии, экспрессируется рецептор, называемый c-kit,
для первого фактора роста — мембранной молекулы клеток
стромы SCF (stem-cell factor) — стволовоклеточного фактора.
Это взаимодействие обеспечивает надлежащее число митозов
еще не поделенных на клоны по рецепторам для антигенов
предшественников В-лимфоцитов.
На следующей стадии — поздней про-В-клетке — экспрес-
сируется рецептор для IL-7, воспринимающий секретируемый
теми же клетками стромы цитокин IL-7. Выявлен и еще один
цитокин, продуцируемый клетками стромы костного мозга,
нокаут гена которого полностью отменяет развитие В-лимфо-
цитов — это PBSF/SDF-1. Данные взаимодействия поддержи-
вают пролиферацию про-В- и больших пре-В-клеток, в ко-
торых уже произошла перестройка генов тяжелой цепи, но
еще не было перестройки генов легкой цепи. Таким образом
накапливаются «полуклоны» В-лимфоцитов с уже известной
специфичностью по тяжелой цепи, но еще неизвестной — по
легкой. Это тоже механизм приумножения разнообразия ан-
тигенсвязывающего репертуара цельных молекул иммуногло-
булинов: с одной и той же тяжелой цепью будут сочетаться
в пары разные варианты легких цепей.
Главные события дифференцировки В-лимфоцитов — пе-
рестройка генов иммуноглобулинов — начинаются на стадии
ранней про-В-клетки с перестройки D-J в генах тяжелых
цепей, причем на обеих гомологичных хромосомах. В поздней
про-В-клетке происходит рекомбинация ДНК V-DJ сначала
на одной из гомологичных хромосом. Если она окажется не-
продуктивной, то та же попытка делается на второй гомоло-
гичной хромосоме. В случае продуктивной перестройки на
первой хромосоме вторая использована не будет.
На следующей стадии в пре-В-клетке происходит пере-
стройка V-J легких цепей, причем сначала одной из цепей —
к или λ, на одной из гомологичных хромосом. Если не полу-
чится продуктивная перестройка с первой попытки в случае
легких цепей, предпринимаются следующие.
Клетки, в которых не получилось ни одной продуктивной
перестройки в генах тяжелых и легких цепей, погибают по ме-
ханизму апоптоза — явления, весьма распространенного для
лимфоцитов.
94

4.7. Рецептор В-лимфоцитов для антигена
Экспрессия на поверхности клетки продуктов перестроенных
генов иммуноглобулинов, кроме того, что является главным
«опорным» параметром конечной цели всей дифференциров-
ки В-лимфоцитов, в динамике служит еще и решающими
ориентирами процесса развития этих клеток.
Собственно связывание антигена — функция вариабельных
доменов димера из тяжелой и легкой цепей иммуноглобули-
на во всех физических состояниях молекулы этого белка, но
чтобы быть рецептором для антигена на клетке «чистой» мо-
лекулы иммуноглобулина мало. Кроме того, что мембранная
форма иммуноглобулина имеет дополнительный гидрофобный
трансмембранный участок полипептида в тяжелых цепях, в
формировании BCR участвуют еще два обязательных полипеп-
тида, называемые (неудачно)
Igtx
(СР79л)а1яв(СР79Ъ)
(рис. 4.6). Дело в том, что
собственный
цитоТВгазШтический
участок трансмембранной формы тяжелых цепей состоит из
остатков всего 3 аминокислот. Этого мало, чтобы иметь эф-
фективные связи с внутриклеточной метаболической «маши-
ной». Рецептор же по определению не только воспринимает
сигнал (физически связывает лиганд), но и проводит его
внутрь клетки. Так вот компоненты BCR
Igcx
и
IgjS
своими
цитоплазматическими участками молекулы связаны с внутри-
клеточными тирозинкиназами, что и обеспечивает проведе-
ние сигнала от связывания антигена внутрь клетки, чтобы та
могла изменить свой метаболизм в соответствии с внешними
запросами. В цитоплазматических участках
Iga
и
Ig/?
присут-
ствуют характерные последовательности остатков аминокис-
V\
A-/
%\
19
7777
\:>л
л</
\
Рис.
4.6. Строение рецептора
В-лимфоцита для антигена
(схема).
95
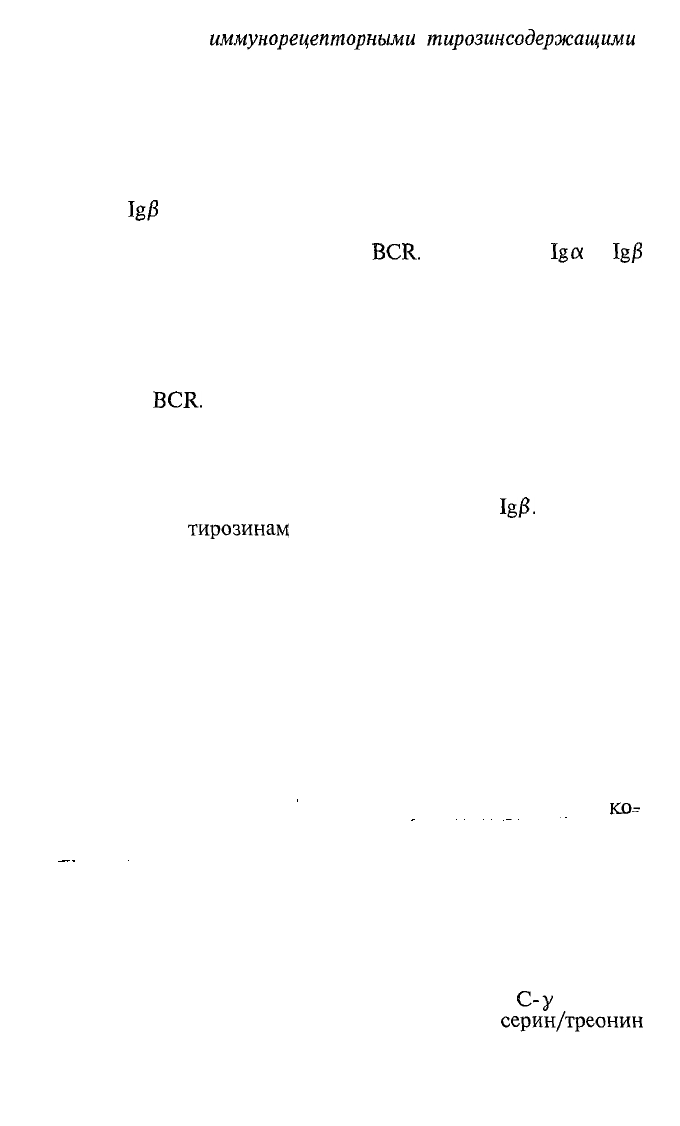
лот, называемые
иммунорецепторными
тирозинсодержащими
активирующими последовательностями (ГТАМ — immunorecep-
tor tyrosine-based activation motifs). Такие же последовательно-
сти присутствуют в проводящих сигнал компонентах рецеп-
тора Т-клеток для антигена. Таким образом известно, что
первой биохимической реакцией активации внутриклеточных
процессов после связывания рецептором антигена является
фосфорилирование остатков тирозина в ITAM.
Iga и
Ig/3
имеют по одному внеклеточному домену, кото-
рым они прочно нековалентно связаны с тяжелыми цепями
иммуноглобулинового компонента
BCR.
Экспрессия
Iga
и
Ig|3
начинается на стадии про-В-клетки и поддерживается в тече-
ние всего онтогенеза В-лимфоцита до самой терминальной
стадии — плазмоцита, на котором экспрессия BCR прогрес-
сивно уменьшается до полного исчезновения.
Для того чтобы произошла эффективная активация В-клет-
ки через BCR, необходима перекрестная сшивка антигеном
нескольких
BCR.
Для этого молекула антигена должна иметь
повторяющиеся эпитопы на своей поверхности. Дальнейшие
события активации В-лимфоцита показаны на рис. 4.7.
Выявлены 4 тирозинкиназы, ассоциированные с BCR: Fyn,
Btk, Lyn и Syk. Сначала первые 3 обеспечивают фосфорили-
рование двух остатков тирозина в ITAM Iga и
Ig/?.
К фосфо-
рилированным
тирозинам;
присоединяется и тем активирует-
ся к действию Syk, продолжающая активационный каскад.
Тирозинкиназы активируются в результате фосфорилирования
в одном месте и ингибируются в результате тоже фосфори-
лирования, но в другом месте молекулы: так устроено, что-
бы процесс активации клетки не принимал характера «враз-
нос».
Для активации необходимо дефосфорилирование ингиби-
ционных участков молекул тирозинкиназ. Такое дефосфори-
лирование катализирует мембранная тирозинспецифичная
фосфатаза CD45. Эта молекула имеет несколько изоформ,
экспрессирована на всех белых клетках крови, поэтому у нее
есть второе название — общий антиген лейкоцитов.
Внутри клетки действует еще одна фосфатаза — SHP,
ко=
торая дефосфорилирует активационные тирозины, чем огра-
ничивает процесс активации лимфоцита. Мыши, у которых
этот фермент отсутствует по причине мутации, реагируют на
существенно меньшие дозы антигенов, чем нормальные
мыши, у них необыкновенно повышен уровень пролиферации
лимфоцитов, и эти мыши умирают через несколько недель
после рождения с клиническими признаками разлитой ауто-
иммунной патологии.
Тирозинкиназа Syk активирует фосфолипазу
С-у
(PLC-y)
и Ras. Ras в свою очередь активирует Raf—
серин/треонин
киназу, которая фосфорилирует внутриклеточные белки по
96

Тирозинкиназа Syk
Тирозинкиназа
Syk
связывается
с фосфорилированными
цитоплазматическими
участками молекул
Iga
и
I
и при этом становится
активной
Тирозинкиназы
Blk,
Fyn, Lyn
Связывание BCR с антиге-
ном вызывает активацию
ассоциированных с рецеп-
тором
тирозинкиназ
Активированные киназы
фосфорилируют по остатку
тирозина
цитоплазматичес-
кие участки молекул
BCR
Активированные киназы
фосфорилируют и тем
"активируют фосфолипазу
С^у
и Ras.
Тирозинфосфатаза CD45
отщепляет ингибирующие
фосфаты от
ассоциирован-
ных с BCR
киназ,
чем
способствует их активации
Фосфолипаза
C-γ
расщепляет фосфатидил-инозитолди -
фосфат
(Р1Р
2
)
с образованием диацилглицерола (DAG) и
инозитолтрифосфата
(1Р
3
)
DAG активирует
протеинкиназу
С
(серин-треонинкиназа).
Фосфорилирование белков
клетки по остаткам серина
и треонина
1Р
3
способствует
повышению концентрации
Са
2+
внутри клетки
Активация
Са
2+
—зависимых
ферментов
Ras активирует серин-
треонинкиназу Raf
Активная Raf фосфорили -
рует белки клетки по остат-
кам серина или треонина
Активация белков, взаимодействующих с ДНК. В результате происходит транскрипция
с определенных генов
Рис.
4.7. Активация В-лимфоцита: внутриклеточная передача «сигнала».
остаткам серина или треонина, что вносит свой вклад в ак-
тивацию ДНК-связывающих белков и тем самым способно
инициировать транскрипцию с генов.
Фосфолипаза
С-у
катализирует расщепление фосфатидил-
инозитола бифосфата на диацилглицерол (DAG) и инозитол
трифосфат
(1Р
3
).
DAG активирует протеинкиназу С — серии/
треонин киназу, которая начинает фосфорилировать белки по
остаткам серина или треонина, что, как и при работе Raf,
заканчивается активацией транскрипции с генов. Инозитол
4-544
97

трифосфат стимулирует повышение в клетке концентрации
свободных ионов Са. В результате активируются кальцийзави-
симые
ферменты, что также действует в направлении акти-
вации транскрипции с генов.
На клеточном уровне активация представляет собой соче-
тание двух феноменов — пролиферации и/или биосинтеза спе-
цифических белков.
О том, что проведение сигнала внутрь клетки — не толь-
ко конечная цель, но и необходимо для самого процесса диф-
ференцировки, свидетельствует тот факт, что генетический
дефект в тирозинкиназе Btk (Bruton's tyrosine kinase) имеет
следствием иммунодефицитную патологию с полным отсут-
ствием у человека В-лимфоцитов — Х-сцепленную агамма-
глобулинемию
Брутона (Bruton's
X-linked
agammaglobuline-
mia — XLA).
4.8. Стадии лимфопоэза В-лимфоцитов
Введение в зародышевые клетки перестроенного трансгена
тяжелой цепи полностью подавляет перестройку одноименных
собственных генов клетки. Но если трансген не содержит кода
для трансмембранного участка тяжелой цепи, то трансген не
мешает перестройке собственного одноименного гена. Следо-
вательно, для того чтобы в конечном счете дифференциро-
ванный
В-лимфоцит
имел строго один вариант тяжелой цепи
и один вариант легкой, еще в процессе дифференцировки
необходима экспрессия тяжелой цепи на мембране. Так оно
и есть. Как только в клетке произошла трансляция полипеп-
тида тяжелой цепи, он экспрессируется на мембране в со-
ставе так называемого пре-В-рецептора. Чтобы это могло слу-
читься, в про-В-клетке синтезируются два специальных по-
липептида, которые нековалентно соединяются друг с другом
и это соединение называют суррогатной легкой цепью. Один из
этих полипептидов —
λ5,
второй — VpreB. Таким образом,
пре-В-клеточный рецептор состоит из
λ
5 + VpreB +
μ-цепь
+
+
Ig«
+
\%β.
Его экспрессия транзиторна, но абсолютно не-
обходима для правильной дифференцировки В-лимфоцитов.
После экспрессии пре-В-рецептора временно инактивируют-
ся белки RAG, и клетки вступают в процесс интенсивной
пролиферации, которая прекращается с исчезновением этого
рецептора. После завершения этой волны пролиферации вновь
экспрессируются
RAG1
и RAG2 и начинается перестройка
генов легкой цепи. Как только это произойдет, на развиваю-
щемся В-лимфоците будет экспрессироваться дефинитивный
BCR состава: L-цепь + μ -цепь +
Ig«
+
Ig/J.
Эту стадию раз-
вития называют незрелым
В-лимфоцитом.
Маркером завершения В-лимфопоэза — образования зре-
лого неиммунного В-лимфоцита, готового к выходу из кост-
98

ΙΙΟΓΟ
мозга в периферическую лимфоидную ткань, — являет-
ся коэкспрессия на мембране двух типов BCR — с IgM и IgD
(задействуется
альтернативный сплайсинг РНК-транскрипта с
μ
δ-гена
тяжелой цепи).
Прежде чем произойдет экспрессия на мембране IgD, но
после того как произошла экспрессия BcR с полноценным
IgM,
в развитии В-лимфоцитов «предусмотрен» существенный
и обязательный этап дифференцировки — селекция (апоптоз)
аутореактивных клонов в местах прохождения лимфопоэза, т.е.
па территории костного мозга. В природе устроено так, что
связывание антигена незрелой В-клеткой, на которой есть
антигенраспознающий рецептор с IgM, но еще нет рецепто-
ра с IgD, является сигналом для апоптоза, т.е. запрограмми-
рованной гибели клетки. Таким образом, из случайного ре-
пертуара по антигенсвязывающим рецепторам на исходе лим-
фопоэза убираются В-лимфоциты, несущие рецепторы, спо-
собные с высокой аффинностью связывать белки собственных
клеток и растворимые белки, присутствующие в достаточных
количествах на территории костного мозга. Такой механизм
толерантности к своему называют делецией клона (clonal
deletion).
Толерантностью в иммунологии называют отсутствие им-
мунного ответа конкретной особи на тот или иной (те или
иные) антигены, на который(ые) другие особи либо та же
особь, но при иных конкретных условиях онтогенеза, потен-
циально способны развивать иммунный ответ. Делеция клона —
не единственный механизм установления толерантности к
конкретному антигену со стороны В-лимфоцитов. Известно
еще два механизма: развитие состояния ареактивности (или
анергии) и
«редакция»
рецептора по антигенной специфично-
сти. Эти два механизма действуют в периферических лимфо-
идных тканях.
Реальность механизма делеции клона, специфичного к
мультивалентному антигену, который экспрессирован на мем-
бранах собственных клеток, хорошо видна в экспериментах с
трансгенными мышами. Например, в таком эксперименте, в
котором трансгены —
гены
легкой и тяжелой цепи молекулы
иммуноглобулина, специфичного к молекулам главного комп-
лекса гистосовместимости I класса
Н-2К
Ь
.
У таких мышей бла-
годаря описанным выше закономерностям все В-лимфоциты
имеют один и тот же BcR с иммуноглобулином, кодируемым
трансгеном. Если мыши-реципиенты такого трансгена в соб-
ственном организме не имеют антигена-мишени (т.е.
Н-2К
Ь
),
у них имеется нормальное количество В-лимфоцитов в пери-
ферических лимфоидных тканях, только все они с одним и
тем же рецептором. Но если мыши-реципиенты сами имеют
ген/антиген
Н-2К\
то у них находят нормальное количество
пре-В-клеток, которые, однако, все погибают на территории
4* 99
костного мозга апоптозом и в периферических лимфоидных
тканях В-лимфоцитов совсем нет. По тому же механизму кло-
нальной делеции погибают и В-лимфоциты на периферии,
если они несут рецептор, способный связывать молекулы
мембран клеток, которые представлены в большом количе-
стве в тех или иных тканях (например, в печени).
Если незрелый В-лимфоцит (с
IgM,
но без
IgD)
связывает
растворимый антиген (например, в организме дважды
транс-
генных мышей: один трансген кодирует синтез растворимого
белка, второй — антитела к нему), то лимфоцит не элими-
нируется апоптозом, а остается в организме, но приобретает
состояние анергии: в результате связывания антигена с рецеп-
тором не наступает активация лимфоцита к иммунному отве-
ту, наоборот, развивается блок проведения сигнала.
У зрелых
В-лимфоцитов
в
периферических
лимфоидных
тканях на большие дозы растворимых антигенов и особен-
но при отсутствии адекватного взаимодействия с Т-хелпе-
рами так же развивается состояние анергии.. Такие клетки
долго не живут и в течение нескольких дней все равно поги-
бают.
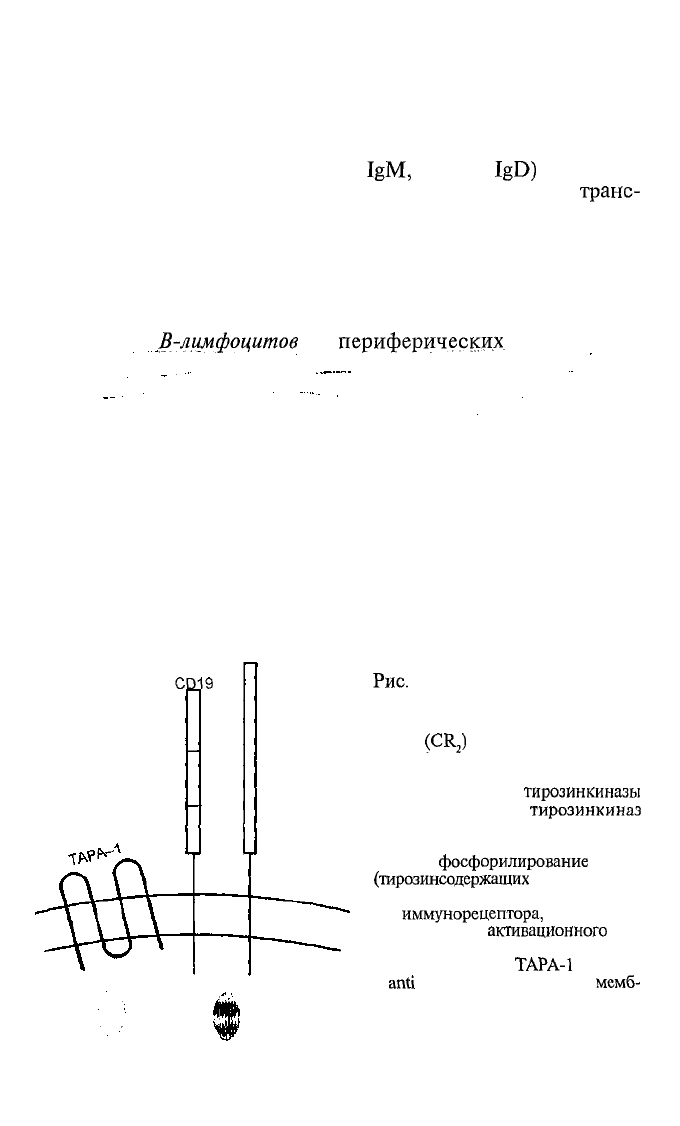
Для полноценной реакции на антиген лимфоциту мало толь-
ко рецептора для антигена. У лимфоцитов есть еще такой обя-
зательный фактор, как корецепторный комплекс мембранных
молекул, связанных с внутриклеточными системами проведе-
ния сигналов. Не на каждом антигене есть повторяющиеся
эпитопы, следовательно, не каждый антиген способен вы-
звать перекрестную сшивку
(или агрегацию) BCR. Вот
CR2
CD19
Scr
PI3
Рис.
4.8. Корецепторный комп-
лекс В-лимфоцита.
Связывание рецептора для компле-
мента
(CR
2
)
с лигандом приводит
к фосфорилированию молекулы
CD19. Фосфорилированная молеку-
ла CD19 связывает
тирозинкиназы
Lyn (из семейства
тирозинкиназ
Scr) и PI-3 (фосфатидилинозитол-
3-киназу), которые вносят свой
вклад в
фосфорилирование
ITAM
(тирозинсодержащих
последователь-
ностей в цитоплазматическом учас-
тке
иммунорецептора,
обеспечива-
ющих передачу
активационного
сиг-
нала) BCR, что усиливает актива-
цию В-лимфоцита. ТАРА-1 (от Target
of
anti
proliferative antibody) —
мемб-
ранная молекула, структурно ассо-
циированная с рецептором В-лим-
фоцита для антигена; ее функции
точно еще не охарактеризованы.
100