Санагурський Д.І. Об’єкти біофізики: монографія
Подождите немного. Документ загружается.

371
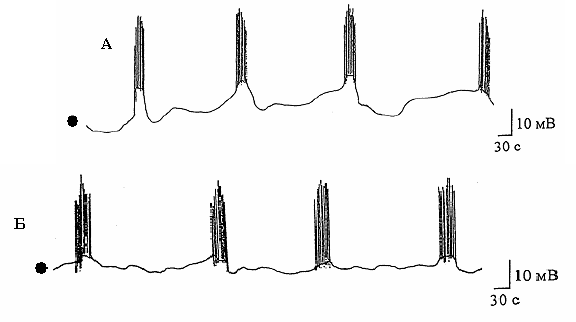
деполяризувалася мембрана в межах 2–3 мВ протягом 5 с. Пауза
між сусідніми пачками триває 150 ± 0,3 с. Тривалість окремої
пачки становить 25–30 с.
Рис. 10.2. Типи електричної активності клітин ектодерми гідри:
А – величина потенціалу спокою -40 мВ;
Б – величина потенціалу спокою -25 мВ.
Примітка. Крапками відмічено середнє значення потенціалу спокою для
кожного із записів
Останній ПД у пачці закінчується невеликою
гіперполяризацією (на 3–5 мВ) мембрани відносно величини
ПС, яка переходить у повільну деполяризацію протягом
міжпачкового інтервалу. Період спокою між пачками імпульсів
становить 100 ± 5 с (n=15). Серії складаються переважно з 7–
10 спайків (рис. 10.3–10.4).
Незначна кількість електричної активності мала вигляд
поодиноких ПД. Отже, при ПД, яким передує повільна
деполяризація (при значеннях МП вищих, ніж –38 ± 0,3 мВ)
можна припустити, що така деполяризація відображає актив-ність
пейсмекерної клітини, а коли ж повільної деполяризації немає,
372
А
Б
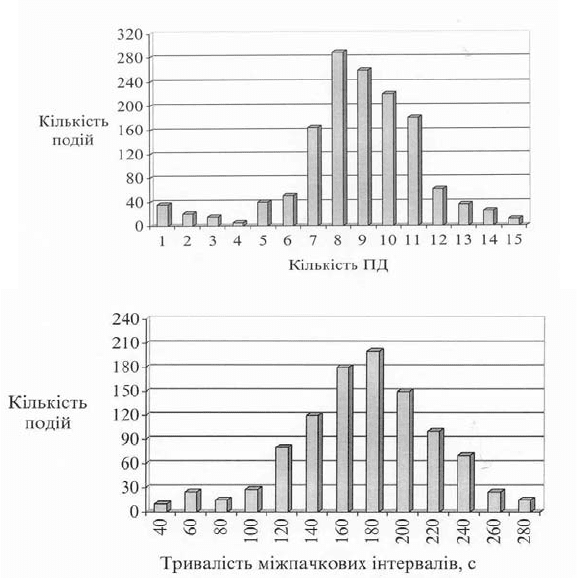
Рис. 10.3. Характеристика пачкової електричної активності клітин
ектодерми гідри у нормі при статистичному обробленні даних (n = 15):
А – кількість ПД у пачці (310 зареєстрованих пачок); Б – тривалість
міжпачкових інтервалів (n = 200)
або вона мало помітна (при значеннях МП нижчих, ніж –38 ± 0,3
мВ), то це означає, що мікроелектрод знаходиться в пейсмекерній
клітині, до якої збудження надходить від пейсмекерної клітини,
що генерує ПД з повільною пейсмекерною деполяризацією.
373
Б
А
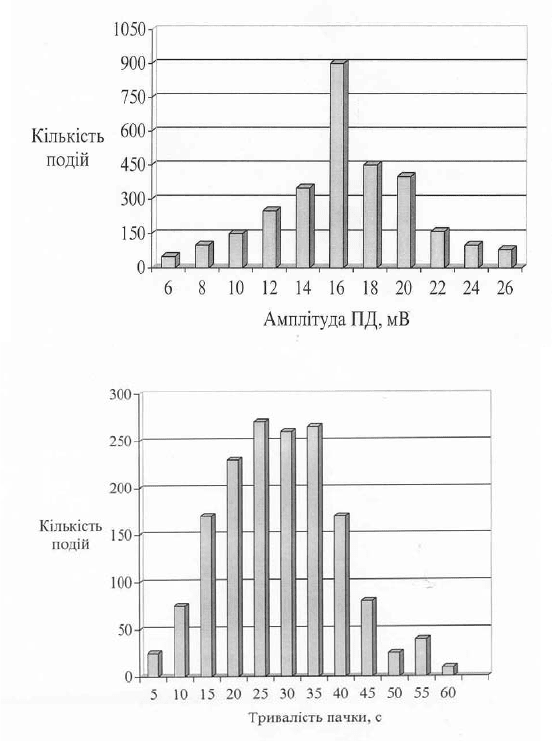
Рис. 10.4. Характеристика пачкової електричної активності клітин
ектодерми гідри у нормі при статистичному обробленні даних (n = 15):
А – амплітуда ПД (900 спайків); Б – тривалість пачки (n = 280)
Аналіз спонтанної електричної активності клітин
ектодерми прісноводної гідри за умов нормального
культурального середовища засвідчив, що поздовжні скорочення
епітеліально-м’язових клітин ектодерми Hydra oligactis мають
374
ступінчасту динаміку. За електричними параметрами це явище
було охарактеризоване як спонтанно активована спайкова
електрична активність (Р.В. Тизьо, 1998), яку також досліджують
на ізольованих клітинах ектодерми Hydra attenuata Pallas
(R.D. Campball et al., 1976).
За проведеним аналізом змін параметрів ПЕА та
порівнюючи їх з даними інших авторів (М. Клевець, 1987;
Л. Страйер, 1985; Р. Тизьо та ін, 1988; М. Шуба, 1965; R. Stampfli,
S. Nishie, 1956), де при різних концентраціях іонів Na, Са
2+
, К
+
використано специфічні блокатори йонних каналів цих катіонів,
можна підтвердити висновок, що ПД клітин ектодерми
генеруються за участю катіонів Na
+
; транспортування йонів Са
2+
спричиняє опосередковану дію на параметри ПЕА; формування
фази реполяризації ПД, а також слідова пачкова гіперполяризація
пов’язані з активуванням К
+
каналів з клітини.
Відомо, що для клітин ектодерми кишковопорожнинних
характерна епітеліальна електрична провідність, яка
координується завдяки нервовим клітинам (J. Itayama, O. Sawada,
1995).
Використовуючи мікроелектродну техніку та метод петч-
клемп встановили, що нервові клітини Hydra attenuata, яким
властивий від’ємний мембранний потенціал спокою (M. Hassen et
al., 1996), здатні спонтанно генерувати серії ПД (R. Campball et al.,
1976; O. Koizumi et al.,1990).
Оскільки дані про електричну активність нервових клітин
ектодерми гідри фрагментарні, доцільно дослідити наявність
ГАМК- та глютаматних рецепторів на клітинах ектодерми
375
прісноводної гідри і з’ясувати роль цих рецепторів у регулюванні
ЕА клітин ектодерми Hydra oligactis.
10.10. Äèíàì³êà åëåêòðè÷íèõ
ïàðàìåòð³â êë³òèí åêòîäåðìè
ïð³ñíîâîäíî¿ ã³äðè ïðè 䳿
áëîêàòîð³â ãàìà-
àì³íîìàñëÿíî¿ êèñëîòè òà
ãëþòàìàòíèõ ðåöåïòîð³â
У процесі вивчення динаміки розвитку
дифузної нервової системи Hydra actinia (J.C. Kinnamon,
J.A. Westfall, 1982) встановлено, що більшість нейронів
зосереджена в ділянці шлунка та підошви (O. Koizumi et al., 1990),
де вони постійно продукуються разом з безперервним поділом
епітеліальних клітин. Схаллер та ін. (H.C. Schaller et al., 1996),
вивчаючи розвиток нервових клітин гідри, виявили компоненти
провідних шляхів цих клітин також у ділянці гіпостома та
підошви. Особливості розвитку нервових клітин гіпостома
досліджували за допомогою пептидів, які є активаторами голови та
низькомолекулярних дієвих речовин – інгібіторів підошви.
Оскільки у ссавців активатори морфогенезу голови продукуються
біля нервових та нейроендокринних клітин, подібність процесів у
нервовій системі гідри (J. Kinnamon, J. Westfall, 1982; O. Koizumi,
H. Bode, 1991; O. Koizumi et al., 1990) та здатність нервових клітин
відповідати на оточуючі сигнали (M. Ohashi et al., 1999; H. Schaller
376
et al., 1996) дає можливість припускати те, що ГАМК– та
глютаматні рецептори наявні на клітинах ектодерми гідри. Метою
наших дальших досліджень стало виявлення рецепторів на
клітинах ектодерми прісноводної гідри мікроелектродним
методом з використанням специфічних блокаторів.
Шляхом біохімічного аналізу гомогенату клітин Hydra
vulgaris було доведено, що в досліджуваних клітинах наявні ГАМК-
рецептори (M. Hassen et al., 1996). Автори зазначають, що вони
чутливі до мусцімолу та бензодіазепінів і беруть участь у
регулюванні харчової реакції. Також досліджено вплив
нейростероїдів, загальних анестетиків та антагоністів ГАМК
А
-
рецепторів (A. Concas et al., 1998; P. Pierobon et al., 1995). Такі
нейростероїди, як тетрагідропрогестерон і тетрагідродезокси-
кортикостерон підвищували зв’язування [3H]-ГАМК з
мембранами гідри. Ліганди діазепаму бензодіазепінового рецеп-
тора (100 мкмоль/л), клоназепаму (100 мкмоль/л) та абекарнілу (30
мкмоль/л) теж підвищували зв’язування [3H]-ГАМК з мембранами
гідри. Цей ефект усувався під впливом флумазенілу, антагоніста
бензодіазепіну (100 мкмоль/л). Навпаки, 4-хлородіазепам, ліганд
периферійних бензодіазепінових рецепторів не впливав на
зв’язування [3H]-ГАМК у мембранах гідри.
Загальні анестетики, пропофол та альфаксалон, подібно
діяли (+38% та +30% відповідно) на зв’язування [3H]-ГАМК.
Більше того, [3H]-ГАМК зв’язування в мембранах гідри повністю
блокувалось антагоністом ГАМК
A
- рецепторів SR 95 531.
Модуляція ГАМК
A
- рецепторів in vitro під впливом різних
препаратів була пов’язана з їхньою дією на харчову реакцію гідри,
377
яку викликали глютатіоном у живих тварин. Тетрагідропро-
гестерон та тетрагідродезоксикортикостерон (від 1 до 10 мкмоль/л)
впливали, залежно від дозування, тривалості відкривання
ротового отвору, зумовленого глютатіоном (10 мкмоль/л) при
максимальних впливах +33% та 29% відповідно. Альфаксалон
(10 мкмоль/л) аналогічно (+33%) збільшував ефект глютатіону.
Ефекти стероїдів на харчову реакцію блокувались SR
95 531, залежно від дозування, t-бутилбіклофосфоротіонатом
(1 мкмоль/л), специфічним блокатором хлорних каналів, але не
бікукуліном.
Ці результати засвідчили, що модуляція ГАМК
А
-
рецепторів стероїдами, яка є характерною для хребетних та
безхребетних тварин, та фармакологічні властивості цих рецеп-
торів збереглися в ході еволюції (A. Concas et al., 1998).
3-гідрокси-5-прегнан-20-один (тетрагідродезоксикортико-
стерон, THDOC) є найбільш потенційними та ефективними і
позитивними алостеричними модуляторами центральних ГАМКA-
рецепторів у ссавців. Діючи в особливо визначеному місці стероїда
в рецепторному комплексі ГАМК
А
, ці складові підсилюють
активність каналів (K.W. Gee, 1988; J.J. Lambert et al., 1995; N.C. Lan
et al., 1991; S.M. Paul, R.H. Purdy, 1992; J.A. Peters et al., 1988). Низькі
наномолярні концентрації цих нейростероїдів посилюють струми,
спричинені ГАМК у нейронах, тоді як вищі концентрації прямо
стимулюють С1
-
-потоки (N.L. Harrison et al., 1987; M.D. Majewska
et al., 1986; J.A. Peters et al., 1988). Уci ці ефекти подібні до таких
ефектів, зумовлених за допомогою класичних позитивних
алостеричних модуляторів ГАМК
А
-рецептора, як бензодіазепіни,
378
барбітурати та загальні анестетики, i є протилежними до ефектів,
зумовлених антагоністами ГАМК (G.A. Cottrell et al., 1987;
T.G. Hales, J.J. Lambert, 1991; R.L. Macdonald, R.W. Olsen, 1994;
J.A. Peters et al., 1988).
Хоча численними дослідженнями доведено наявність
ГАМК-рецепторів у нервовій системі хребетних та безхребетних, а
також у бактерій, чутливість цих рецепторів до стероїдних
анестетиків з’являлась у процесі еволюції (D. Belelli et al., 1996; R.
Chen et al., 1994; A. Hosie, D. Sattelle, 1996; G. Lunt, 1991; J. Schaeffer,
A. Bergstrom, 1988). Отже, якщо стероїдна модуляція ГАМК
А
-
рецепторів добре збереглася в процесі еволюції хребетних, ГАМК
А
-
рецептори у безхребетних мають обмежену чутливість або
нечутливість до статевих стероїдів (A.M. Hosie, D.B. Sattelle, 1996;
A.E. Oliver et al., 1991; J.J. Rauh, 1993).
Досліджено також вплив нейростероїдів, загальних
анестетиків, лігандів бензодіазепінового рецептора та ГАМК
антагоністів, які є модуляторами ГАМКА-рецепторів у ссавців, на
ГАМК-рецептори, наявні в мембранах клітин гідри. Більше того,
було досліджено зв’язок між біохімічними впливами цих речовин
на ГАМКА-рецептори гідри та модуляцію її харчових реакцій.
Поведінкова реакція спричинена малою кількістю глютатіону
(GSH) після того, як здобич було впіймано або за допомогою GSH,
якщо здобичі не було, вважається простою та надійною моделлю
для дослідження хеморецепції (H. Grundfest et al., 1955; M. Kim et
al., 2000; W. Loomis, 1969).
Харчову реакцію вивчали за допомогою процедур,
описаних Ленгофом (H.M. Lenhoff et al., 1983). Інкубаційний час
379
зв’язування [3H]-ГАМК при температурі 4
о
С стабілізувався через
п’ять хвилин. Попередньо було з’ясовано (J.A. Westfall, D.R. Argast,
1982), що дослідження насиченості зв’язування [3H]-ГАМК на
заморожування-розморожування, а також на відмивання
мембранних препарувань з гідри дали змогу виявити наявність
лише однієї популяції місць зв’язування, що засвідчує низьку
здатність (В
max
= 4,75 пМ/мг білка) та високу спорідненість (КD =
76 пМ). Як і очікували, бікукулін (1–100 мкмоль/л) не дуже
вплинув на зв’язування [3H]-ГAMK у гомогенатах клітин гідри, але
(залежно від дозування) мав інгібувальний вплив на зв’язування з
мембран мозку щура (табл. 10.3).
Таблиця 10.3
Дія бікукуліну та SR 955 331 на зв’язування [3H]-ГАМК у
гомогенаті клітин Hydra vulgaris
Речовина
Концентрація
(мкмоль/л)
Специфічне
[3H]-ГАМК
%
Бікукулін 1 97.2 ± 6.0 97
Бікукулін 10 91.3 ±8.0 91
SR 955 331 1 27.8 ± 2.0 27
SR 955 331 10 2.0 ±0.3 2
Щоб встановити особливості [3H]-ГАМК зв’язування було
досліджено впливи антагоніста SR 955 331 ГАМК
А
-рецептора,
похідного ГАМК піридазинілу (M. Heauime et al., 1986). На
противагу, бікукулін SR 955 331 (1-10 мкмоль/л) інгібував [3H]-
ГАМК зв’язування у гомогенаті клітин гідри, і з кори головного
мозку щура, залежно від концентрації табл. 10.3).
Характеризуючи біохімічні особливості ГАМК-
передавання в гідри, оцінювали вплив центральних та
380
периферійних лігандів бензодіазепінового рецептора (діазепам,
клоназепам, абекарніл, 4-хлородіазепам) та загальних анестетиків
(пропофол, альфаксалон) на [3H]-ГАМК зв’язування з гомогенату
клітин гідри. Діазепам (100 мкмоль/л), клоназепам (100 мкмоль/л),
та бетакарболін, похідний абекарнілу (30 мкмоль/л) збільшував
[3H]-ГАМК зв’язування подібно (максимальний вплив +22, +20,
+24 %, відповідно), тоді як збільшення, спричинені меншими
дозами (3–10 мкмоль/л) названих препаратів, не досягли
статистичної достовірності.
Більші концентрації цих препаратів не збільшували [3H]-
ГАМК зв’язування. Вплив діазепаму, клоназепаму та абекарнілу
був антагонізований флумазенілом (100 мкмоль/л) центрального
бензодіазепінового рецептора (табл. 10.4). Як встановлено
попередньо (G. Biggio et al., 1990; M. Serra et al., 1992) діазепам,
клоназепам та абекарніл також збільшили [3H]-ГАМК зв’язування
з мембранами клітин мозку щура. Навпаки, 4-хлородіазепам (3–
30 мкмоль/л) не підтвердив [3H]-ГАМК зв’язування ні з
мембранами клітин гідри, ні з мембранами нейронів щура.
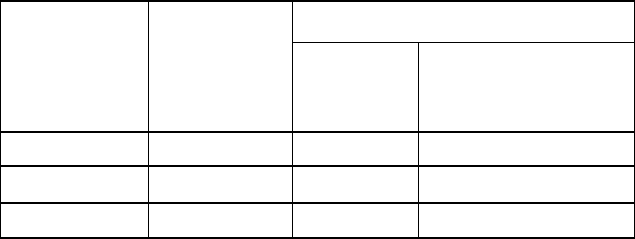
Таблиця 10.4
Дія лігандів бензодіазепінового рецептора за наявності
антагоніста флумазенілу (100 мкмоль/л) на зв'язування [
3
H]-ГAMK
у гомогенаті клітин гідри Hydra vulgaris
[
3
H]-ГАМК зв'язування (%)
Речовина
Концентрація
(мкмоль/л)
Речовина
Флумазеніл
+
Речовина
Діазепам 100 124,2 ± 4.0 106,0 ± 3.0
Клоназепам 100 21,0 ±5 .0 102,0 ± 4.0
Абекарніл 30 126,0 ± 6.0 98,0 ± 7.0