Реферат - Генетика популяции и эволюция
Подождите немного. Документ загружается.

Министерство образования и науки
Воронежский государственный университет
Биолого-почвенный факультет
Генетика популяции и эволюция
реферат
Преподаватель Лавлинский А.В.
Студент Зеленина Е.А.
Воронеж 2011
2
СОЖЕРЖАНИЕ
Видообразование 3
Способы видообразования 5
Роль полиплодии в видообразовании 9
Хромосомные перестройки и видообразование 12
Модификации гетерохроматина и видообразование 14
Роль множественных геномных перестроек в видообразовании 16
Изолирующие механизмы 19
Эволюционная дивергенция 24
Динамика популяций 26
Литература 29
3
ВИДООБРАЗОВАНИЕ
Вид – это совокупность особей, сходных между собой, населяющих
определенный ареал, способных скрещиваться между собой, давать
плодовитое потомство, похожее на родителей и отличающихся по ряду
существенных признаков от других подобных совокупностей.
Видообразова́ние;— процесс возникновения новых видов. При этом
генетическая несовместимость новообразованных видов, т.е. их
неспособность производить плодотворное потомство или вообще потомство
при скрещивании называется межвидовым барьером, или барьером
межвидовой совместимости.
Существуют разнообразные теории, объясняющие механизмы
видообразования, ни одна из которых не считается общепризнанной и
полностью доказанной. Одна из причин этого — сложность эмпирической
проверки из-за долговременности изучаемого процесса.
Видообразование – это качественный этап эволюционного процесса.
Это означает, что образованием видов завершается микроэволюция и
начинается макроэволюция.
Микроэволюцию можно рассматривать как эволюцию популяций
–;открытых генетических систем, способных обмениваться генетическим
материалом, а макроэволюцию – как эволюцию совершенных таксонов
–;закрытых генетических систем, которые не способны обмениваться генами
в естественных условиях. Вид занимает промежуточное положение между
открытыми и закрытыми генетическими системами, поэтому биологический
вид можно рассматривать как;устойчивую генетическую систему,
эволюционирующую относительно независимо от других подобных систем.
В то же время биологический вид представляет собой основную
единицу сообщества, поскольку именно виду соответствует собственная
экологическая ниша.
4
Таким образом, для понимания сущности видообразования необходимо
рассмотреть его популяционно-генетические и экологические аспекты.
Видообразование – это сложный, не изученный до конца процесс.
Известно множество механизмов образования новых видов. Но в любом
случае новый вид должен отличаться от материнского или сестринского вида
хотя бы некоторыми наследуемыми признаками и, следовательно, хоты бы
некоторыми аллелями. Таким образом, в ходе видообразования исходный
набор аллелей должен быть замещен новым набором аллелей.
Согласно общепринятой точке зрения, в основе видообразования лежит
принцип дивергенции. В результате дивергенции увеличивается число видов.
Дивергенция (от лат.;divergo;– отклоняюсь, отхожу) – это расхождение
признаков организмов в ходе эволюции разных групп (филетических линий),
возникших от одного предка. Дивергенцией называют также разделение в
процессе эволюции единого таксона на два или несколько. Термин
«дивергенция признаков» введён Ч. Дарвином (1859).
В больших стационарных популяциях (мегапопуляциях)
видообразование затруднено. Например, в средней полосе европейской части
России многие роды высших растений и крупных животных представлены
или одним видом (ель, сосна, дуб, лещина; липа, медведь, волк, лиса,
белка…), или немногими видами (березы пушистая и бородавчатая; заяц-
беляк и заяц-русак…). Это связано с тем, что при большой и постоянной
численности популяций элементарные эволюционные факторы практически
не действуют.
Таким образом, для начала видообразования исходная большая
популяция (мегапопуляция) должна быть разделена на множество
малочисленных изолированных популяций (дробление крупных популяций
на множество мелких, или островных популяций
называется;инсуляризацией). Такая изоляция является первичной и носит
случайный характер.

5
СПОСОБЫ ВИДООБРАЗОВАНИЯ
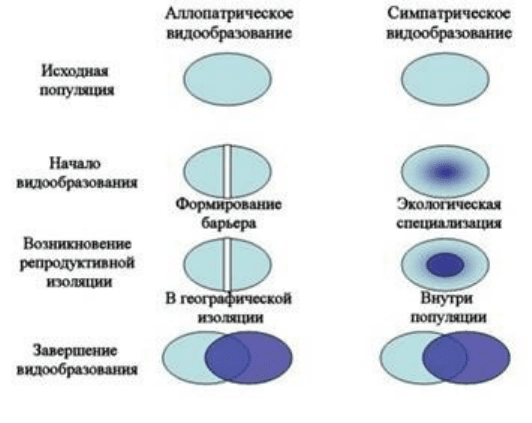
Симпатрическое (экологическое) видообразование
Связано с расхождением групп особей одного вида и обитающих на
одном ареале по экологическим признакам. При этом особи с
промежуточными характеристиками оказываются менее приспособленными.
Расходящиеся группы формируют новые виды. Симпатрическое
видообразование может протекать несколькими способами. Один из них —
возникновение новых видов при быстром изменении кариотипа путём
полиплоидизации. Известны группы близких видов, обычно растений, с
кратным числом хромосом. Другой способ симпатрического
видообразования — гибридизация с последующим удвоением числа
хромосом. Сейчас известно немало видов, гибридогенное происхождение и
характер генома которых может считаться экспериментально доказанным.
Третий способ симпатрического видообразования — возникновение
репродуктивной изоляции особей внутри первоначально единой популяции в
результате фрагментации или слияния хромосом и других хромосомных
перестроек. Особенностью симпатрического пути видообразования является
то, что он приводит к возникновению новых видов, всегда морфологически
близких к исходному виду. Лишь в случае гибридогенного возникновения
видов появляется новая видовая форма, отличная от каждой из родительских.
Особый случай симпатрического видообразования наблюдается в
случае;кольцевых клин, если популяции последовательно друг за другом
заселяют какую-либо географическую область, замыкая кольцо. Краевые
популяции могут при этом оказаться генетически настолько различными, что
будут неспособны образовывать плодовитое потомство.

6
Рис.1. Пример кольцевой клины. Подвиды 1 и 7, возможно, не смогут
скрещиваться между собой
Аллопатрическое (географическое) видообразование
Вызывается разделением ареала вида на несколько изолированных
частей. При этом на каждую такую часть отбор может действовать по-
разному, а эффекты дрейфа генов и мутационного процесса будут явно
отличаться. Тогда со временем в изолировнных частях будут накапливаться
новые генотипы и фенотипы. Особи в разных частях ранее единого ареала
могут изменить свою экологическую нишу. При таких исторических
процессах степень расхождения групп может достигнуть видового уровня.
Рис.2. Наступление моря может привести к изоляции участков суши и популяций,
обитающих на них, что является одной из причин образования новых видов
7
Рис.3. Основные этапы;аллопатрического;и;симпатрического видообразования
Гибридогенное видообразование
При скрещивании различных видов потомство обычно бывает
стерильным. Это связано с тем, что число хромосом у разных видов
различно. Несходные хромосомы не могут нормально сходиться в пары в
процессе мейоза, и образующиеся половые клетки не получают нормального
набора хромосом. Однако, если у такого гибрида происходит геномная
мутация, вызывающая удвоение числа хромосом, то мейоз протекает
нормально и дает нормальные половые клетки. При этом гибридная форма
приобретает способность к размножению и утрачивает возможность
скрещивания с родительскими формами. Кроме того, межвидовые гибриды
растений могут размножатся вегетативным путем.
Существующие в природе естественные ряды гибридных видов
растений возникли, вероятно, именно таким путем. Так, известны виды
пшеницы с 14, 28 и 42 хромосомами, виды роз с 14, 28, 42 и 56 хромосомами
и виды фиалок с числом хромосом, кратным 6 в интервале от 12 до 54. По
некоторым данным, гибридогенное происхождение имеют не менее трети
всех видов цветковых растений.
Гибридогенное происхождение доказано и для некоторых видов
животных, в частности, скальных ящериц, земноводных и рыб. Некоторые
8
виды кавказских ящериц, имеющих гибридогенное происхождение,
триплоидны и размножаются с помощью партеногенеза.
Квантовое видообразование
Квантовое видообразование представляет собой отпочковывание
нового дочернего вида от небольшого периферического изолята обширного
полиморфного предкового вида. Этот процесс впервые описан Майром
(Mayr, 1954*), а в дальнейшем его разработкой занимались Майр (Mayr,
1963*) и другие исследователи.
Термин «квантовое видообразование» предложил для обозначения
этого процесса Грант (Grant, 1963). При географическом видообразовании
обычно наблюдается следующая последовательность: локальная раса—
географическая раса—аллопатрический полувид—вид. В отличие от этого
при квантовом видообразовании имеет место непосредственный переход от
локальной расы к новому виду. Квантовое видообразование представляет
собой, таким образом, сокращенный способ формирования видов.
Этапы видообразования
Процесс видообразования слагается из ряда последовательных этапов:
1) создание изменчивости по множественным генам; 2) образование нового
сочетания аллелей; 3) закрепление этого нового сочетания аллелей в
производной популяции; 4) защита нового сочетания аллелей механизмами
репродуктивной изоляции.
РОЛЬ ПОЛИПЛОДИИ В ВИДООБРАЗОВАНИИ
9
Полиплоидия — увеличение числа хромосом, кратное гаплоидному
набору. В соответствии с этим у растений различают триплоиды (3n),
тетраплоиды (4n) и т. д. У растений новые виды достаточно легко могут
образовываться с помощью полиплоидии — мутации удвоения хромосом.
Возникшая таким образом новая форма будет репродуктивно изолирована
от родительского вида, но благодаря самооплодотворению сможет оставить
потомство. Для животных такой способ видообразования неосуществим,
так как они не способны к самооплодотворению.
Среди растений есть немало примеров близкородственных видов,
отличающихся друг от друга кратным числом хромосом, что указывает на
их происхождение путем полиплоидии. Так, у картофеля, есть виды с
числом хромосом, равным 12, 24, 48 и 72; у пшениц — с 14, 28 и 42
хромосомами.
Полиплоиды обычно устойчивы к неблагоприятным
воздействиям, и в экстремальных условиях естественный отбор будет
благоприятствовать их возникновению. Так, на Шпицбергене и Новой
Земле около 80% видов высших растений представлены полиплоидными
формами.
У растений встречается и другой, более редкий способ
хромосомного видообразования — путем гибридизации с последующей
полиплоидией. Близкородственные виды часто различаются своими
хромосомными наборами, и гибриды между ними получаются
бесплодными вследствие нарушения процесса созревания половых клеток.
Гибридные растения, тем не менее, могут существовать довольно
продолжительное время, размножаясь вегетативно.
Мутация полиплоидии «возвращает» гибридам способность к
половому размножению. Именно таким образом — путем гибридизации
терна и алычи с последующей полиплоидией — возникла культурная слива.
10
На основании данных сравнительной геномики было построено раннее
филогенетическое древо геномных дупликаций, которые произошли в
процессе эволюции покрытосеменных растений (Bowers et al., 2003; Paterson
et al., 2004).
Способность к скрещиванию у покрытосеменных растений из разной
степени дивергировавших групп давали гибридные комбинации, которые
после очередной дупликации геномов могли стать родоначальниками новых
видов. Считают, что вследствие таких событий около 11,4 млн лет назад
появилась кукуруза Z. mays (2n = 20) - древний сегментный аллотетраплоид
(Gaut, Doebley, 1997). Другой пример образования аллотетраплоидов
относится к видообразованию у хлопчатника Gossypium (Adams et al, 2003).
Формирование многих полиплоидных таксонов у покрытосеменных
растений происходило многократно, о чем свидетельствует наличие
полиплоидных рядов и у однодольных и у двудольных (Stebbins, 1971; Grant,
1981). Так, появление гексаплоидной (мягкой) пшеницы T. aestivum (2n = 42;
BBAADD) обусловлено двумя независимыми, сравнительно недавними
событиями межвидовой гибридизации, завершившимися дупликацией и
диплоидизацией геномов около 9,5 тыс. лет назад (Levy, Feldman, 2002).
Результаты внутривидовой гибридизации с последующим удвоением числа
хромосом привели к образованию большого многообразия
автополиплоидных видов, а межвидовой - аллополиплоидных. Среди
покрытосеменных растений, по разным оценкам, насчитывают от 30 до 80 %
полиплоидных видов (Soltis et al, 2003).
Полиплоиды описаны у псилотовидных, моховидных, плауновидных,
хвощевидных, папоротниковидных (Stebbins, 1971; Жизнь растений, 1978). У
папоротниковидных примерно 95 % растений - полиплоиды (Grant, 1981).
Приведенные примеры показывают, что один вид может быть представлен
как несколькими полиплоидными цитотипами, так и анеуплоидами,
сформировавшимися в результате цитогенетической нестабильности