Наумов А.Д. Двустворчатые моллюски Белого моря. Опыт эколого-фаунистического анализа
Подождите немного. Документ загружается.


Глава 10. СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
200
Подкласс
AUTOBRANCHIA
Grobben, 1894
Отряд MYTILIFORMES Fèrussac, 1822
Семейство CRENELLIDAE Gray, 1840
Род CRENELLA Brown, 1827
Crenella decussata (Montagu, 1808)
Mytilus decussatus: Montagu, 1808: 69
Crenella decussata decussata: Скарлато, 1981: 220, рис. 118; Кафанов, 1991: 21
Crenella decussata: G. O. Sars, 1878: 31, tab. 3, fig. 4; Герценштейн, 1885: 646; Дерюгин,
1928: 298; Месяцев, 1931: 68; Филатова, 1948: 430, табл. 108, рис. 11; Ушаков, 1953: 262;
Bernard, 1979: 24, fig. 32, 33; Скарлато, 1981: 220, рис. 118; Bernard, 1983: 20; Наумов и
др., 1987: 220, рис. 130; Coan et al., 2000: 166, pl. 24; Galkin, Voronkov, 2001: 111
Диагноз (атлас, таблица II, 1).
Раковина маленькая, выпуклая, перламутровая, округлая, светло-коричневая, блестя-
щая, равностворчатая, равносторонняя.
J
1
= 1.63 + 0.05; J
3
= 0.625 + 0.004 (Наумов и др., 1987). l = 0.43 + 0.69h; h = 0.56 + 1.25b
(Наумов, Федяков, 1985). Максимальные размеры в Белом море: 4.8 × 5.6 × 3.2 мм (Нау-
мов и др., 1987).
Макушки прозогирные.
Периостракум прозрачный, матовый. Поверхность раковины покрыта тонкими ради-
альными ребрышками.
Лунка и щиток не ограничены.
Лигамент наружный, короткий, погруженный.
Замок дизодонтный. Настоящие зубы отсутствуют, но края
раковины и области маку-
шек несут мелкие зубчики.
Мантийная линия без синуса.
Отпечатки аддукторов почти равны по величине.
Палеонтологические находки. Известен с плиоцена (Bernard, 1983; Coan et
al., 2000), в отложениях Белого моря с – бореали (Невесский и др., 1977; Говберг,
1968, 1970, 1973, 1975).
Распространение. Широко распространенный бореально-арктический вид.
Встречен в Баренцевом, Белом и Карском морях, у берегов Гренландии, Исландии,
Шпицбергена и Земли Франца-Иосифа. В Атлантическом океане расселяется на юг
до Англии и мыса Хаттерас, в Тихом
океане – до Сахалина и Сан-Диего.
По материалам БентБоМ (встречен на 104 станциях), данным К. М. Дерюгина
(1928) (встречен на 8 станциях) и коллекций ЗИН РАН (25 пробы) в Белом море на
мелководьях встречается повсеместно. Кандалакшского желоба с его относительно
большими глубинами избегает. По данным Л. А. Кудерского (1961) обнаружен в
районе Соловецких островов и
в центральной части Онежского залива, но не в его
куту. Это неплохо согласуется с нашими данными, по которым наиболее часто этот
вид встречается в северной части залива, а в кутовой относительно редок (см. при-
ложение 1, 7).
Основные экологические особенности. Стенотермный умеренно тепло-
водный, относительно стенобатный, относительно эвритопный, предпочитающий
илистые и песчаные
грунты вид (Федяков, 1986). По БентБоМ отмечен на глубинах
от 2 до 75 м при температуре от –0.8 до 14.4°С и солености от 23.6 до 28.7‰, в ос-
новном на илистых (34% находок), песчаных (21% находок) и смешанных (33% на-
ходок) грунтах. Средняя биомасса этого вида на тех станциях, где он обнаружен,
составляет 0.597 ± 0.123 г/м
2
, а средняя плотность поселения – 112.3 ± 18.7 экз./м
2
.
По материалам БентБоМ максимальные биомасса (4.016 г/м
2
) и плотность посе-
ления (304 экз./м
2
) отмечены в Западной Соловецкой салме на глубине 20 м в биоце-
нозе Modiolus modiolus на песчаном грунте при температуре 8.4°C (табл. 36).
CRENELLA DECUSSATA
201
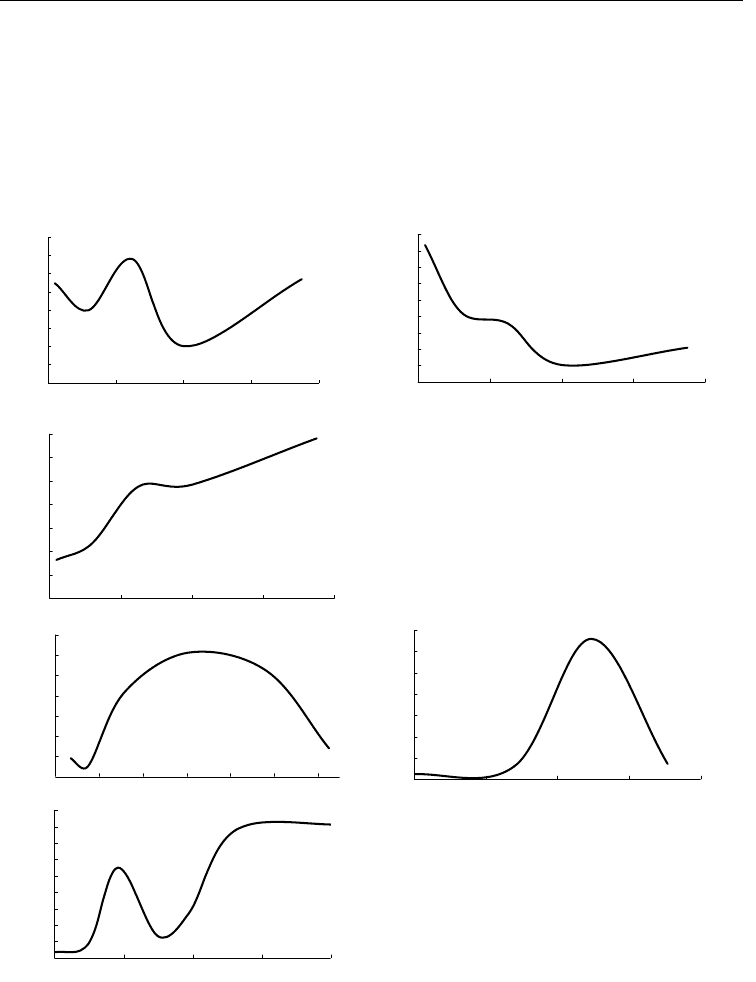
По материалам БентБоМ наибольшие значения биомассы наблюдаются на глу-
бине около 30 м, а плотность поселения довольно равномерно падает с глубиной.
При этом чем глубже были взяты пробы, тем в среднем крупнее оказываются экзем-
пляры этого вида (рис. 100, А, Б, В).
Предпочитает умеренные температуры, поэтому на глубины с высокой солено-
стью не
заходит. Максимальная биомасса встречена на песках, но встречается и на
более крупных осадках – гальке и камнях, скорее всего, из-за того, что в них всегда
есть доля песка. Минимальная биомасса обнаружена на гравийных грунтах
(рис. 101, А, Б, В).
Фильтрующий сестонофаг.
Зрелые ооциты диаметром 120÷200 мкм отмечены с ноября по июнь. Шарообраз
-
ные прикрепленные к грунту кладки встречены в июне. Подвижная трохофора дер-
жится в придонном слое воды. Зародышевая раковина имеет в длину около 600 мкм
(Наумов др., 1987).
Продолжительность жизни в Белом море – 7 лет (Русанова, 1963).
А
0,000
0,100
0,200
0,300
0,400
0,500
0,600
0,700
0,800
0 20406080
Б
0
20
40
60
80
100
120
140
160
180
0 20406080
В
0,000
0,002
0,004
0,006
0,008
0,010
0,012
0,014
0 20406080
А
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
-11357911
Б
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
24 25 26 27 28
В
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
0,01 0,10 1,00 10,00 100,00
Рис. 100. Зависимость биомассы (
А
),
плотности поселения (Б) и среднего веса
экземпляра (В) Crenella decussata от
глубины.
Обозначения см. на рис. 92
Fig. 100. Biomass (А), density (Б) and
mean weight of a specimen (В) in Crenella
decussata plotted against depth.
For legend see fig. 92
Рис. 101. Зависимость биомассы Crenella de-
cussata от температуры (А), солености (Б) и
медианнного размера частиц грунта (В).
Обозначения см. на рис. 94
Рис. 101. Dependence of biomass on temperature
(А), salinity (Б) and medium sediment particle
dimension (В) in Crenella decussata.
For le
g
end see fi
g
. 94

Глава 10. СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
202
Семейство MYTILIDAE Rafinesque, 1815
Подсемейство MUSCULINAE Iredale, 1939
Род MUSCULUS Röding, 1798
Musculus discors (Linnaeus, 1767)
Mytilus discors: Linnaeus, 1767: 1159
Modiolaria discors: G. O. Sars. 1878:29; Дерюгин, 1928: 297; Месяцев, 1931: 67;
Горбунов, 1952: 232, таб. I, фиг. 1, 2
1
Musculus (Musculus) discors: Bernard, 1979:27, fig. 36, 39; Bernard, 1983; Кафанов, 1991: 25
Musculus discors: Филатова, 1948: 429, табл. 108, рис. 8; Скарлато, 1981: 225, рис. 122,
Наумов и др., 1987: 222, рис. 131; Наумов, Федяков, 1994: 56; Coan et al., 2000: 168, pl. 25;
Galkin, Voronkov, 2001: 111
Диагноз (атлас, таблица II, 2).
Раковина вытянутая, темно-оливковая или коричневая, равностворчатая, неравносто-
ронняя, перламутровая.
J
1
=0.672 ± 0.005; J
3
=0.689 ± 0.007 (Наумов и др., 1987). l= –0.01 + 1.54h; h = 1.12 + 1.30b
(Наумов, Федяков, 1985). Аллометрическая зависимость веса тела от длины раковины
W = 0.00027L
2.69
(Максимович, 1978, б). Максимальные размеры в Белом море:
31.1 × 15.8 × 11.6 мм (Наумов и др., 1987).
Макушки прозогирные, смещены вперед.
Периостракум блестящий. Поверхность створок поделена на три поля: переднее, сред-
нее и заднее. Переднее поле равномерно покрыто радиальными ребрышками, среднее –
гладкое; заднее, отделенное от среднего складкой, гладкое или покрыто радиальной исчер-
ченностью.
Лунка
и щиток не ограничены.
Лигамент наружный, погруженный.
Зубы замка отсутствуют. Края створок в области переднего и заднего полей зазубрены.
Мантийная линия без синуса.
Отпечаток заднего мускула-замыкателя округлый, переднего — вытянут вдоль края ра-
ковины. Передний аддуктор меньше заднего и сильно сдвинут к переднему краю створок.
Палеонтологические находки. Известен с плейстоцена (Bernard, 1983) или
даже с плиоцена (Coan et al., 2000). В отложениях Белого моря не отмечен.
Распространение. Широко распространенный бореально-арктический циркум-
полярный вид. Встречен во всех северных морях России, море Бофорта, у берегов
Гренландии. В Атлантическом океане расселяется на юг до Мадейры и о-вов Зелено-
го Мыса и до пролива
Лонг-Айленд, в Тихом океане – до зал. Петра Великого и зал.
Пьюджет-Саунд.
У К. М. Дерюгина (1928), как и большинство других мелководных видов, не от-
мечен. По материалам БентБоМ (встречен на 23 станциях) и коллекций ЗИН РАН (20
проб) в Белом море встречается в Кандалакшском, Онежском, Двинском и Мезен-
ском заливах, а
также на границе Горла и Бассейна (см. приложение 1, 8). То обстоя-
тельство, что в Горле этот вид до сих пор не обнаружен, возможно, указывает на ра-
зобщенность его поселений во внутренних частях моря и в Мезенском заливе.
Основные экологические особенности. Стенотермный умеренно тепло-
водный, относительно эврибатный, стенотопный, обитающий на талломах макрофи-
тов вид (Федяков, 1986). По материалам БентБоМ отмечен на глубинах от 0 до 98 м
при температуре от –1.2 до 13.2°С и солености от 25.4 до 29.0‰, в основном на или-
1
Эти рисунки иллюстрируют разницу скульптуры периостракума заднего поля раковины у
двух подвидов этого вида.
MUSCULUS DISCORS
203
стых (23% находок), каменистых (38% находок) и смешанных (23% находок) грун-
тах. Средняя биомасса этого вида на тех станциях, где он обнаружен, составляет
2.721 ± 1.231 г/м
2
, а средняя плотность поселения – 89.7 ± 31.2 экз./м
2
.
Встречается на слоевищах видов рода Fucus, реже в ризоидах Laminaria, иногда
на нитчатках или непосредственно на грунте (Русанова, 1963).
По материалам БентБоМ максимальные биомасса (1.720 г/м
2
) и плотность посе-
ления (360 экз./м
2
) встречены у мыса Никодимского на глубине 5 м на каменистом
грунте при температуре 4.5°C солености 26.9‰ в биоценозе Phycodrys (табл. 37).
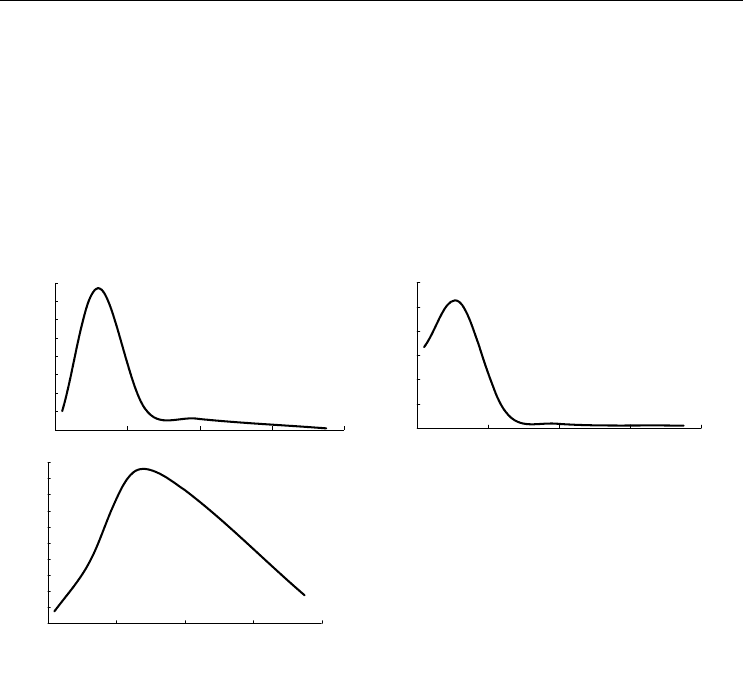
По материалам БентБоМ наибольшие значения биомассы и плотности поселения
наблюдаются на глубинах порядка 10-20 м, а наиболее крупные в среднем экземпля-
ры были обнаружены на глубине 20-40 м (рис. 102).
В Онежском заливе при обследовании 8 экз.
этого вида, в мантийной полости 5 из
них был обнаружен симбиотический бокоплав Metopa glacialis (Krøyer) (Марченков,
1997).
Фильтрующий сестонофаг. Прикрепляется к талломам, часто деформируя слое-
вище, и ризоидам макрофитов биссусом, из нитей которого строит гнездо, в которое
самки откладывают кладки в виде белых закрученных шнуров (Матвеева, 1975,
1979). Может встречаться и на листьях Zostera marina (Rasmussen, 1973). Половой
зрелости
достигает на 1–2 году жизни.
Нерест растянут. Максимум его приходится на летние месяцы. Слизистые кладки
с развивающимися яйцами в гнездах моллюсков встречаются во все сезоны года.
Диаметр зрелых яиц 300÷360 мкм (Кауфман, 1977) или 250 мкм (Матвеева, 1979).
Развитие без пелагической личинки (Матвеева, 1975, 1979), зародышевая раковина
имеет длину около 500 мкм (Наумов и др., 1987).
Продолжительность жизни в
Белом море – 3÷4 года (Максимович, 1980).
А
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0 20406080
Б
0
20
40
60
80
100
120
0 20406080
В
0,000
0,002
0,004
0,006
0,008
0,010
0,012
0,014
0,016
0,018
0,020
0 20406080
Рис. 102. Зависимость биомассы (А),
плотности поселения (Б) и
среднего веса
экземпляра (В)
Musculus discors от
глубины.
Обозначения см. на рис. 92
Fig. 102. Biomass (А), density (Б) and
mean weight of a specimen (В) in
Musculus
discors
plotted against depth.
For legend see fig. 92

глава 10. СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
204
Musculus corrugatus (Stimpson, 1851)
Mytilus corrugatus: Stimpson, 1851: 12
Modiolaria corrugata: G. O. Sars, 1878: 30, tab. 19, fig. 2; Герценштейн, 1885: 645;
Knipowitsch, 1902: 399; Дерюгин, 1928: 297; Месяцев, 1931: 66; Горбунов, 1952: 235, таб. I,
фиг. 4, 5
Musculus (Musculus) corrugatus: Bernard, 1979: 26, fig. 35; Кафанов, 1991: 25
Musculus corrugatus: Филатова, 1948: 429, табл. 108, рис. 6; Ушаков, 1953: 262; Lubinsky,
1980: 25, pl. 4, fig. 7-10; Скарлато, 1981: 227, рис. 123; Наумов и др., 1987: 222, рис. 132;
Наумов, Федяков, 1994: 56; Galkin, Voronkov, 2001: 111
Диагноз (атлас, таблица II, 3).
Раковина вытянутая, темно-оливковая или коричневая, равностворчатая, неравносто-
ронняя, перламутровая.
J
1
=0.708 ± 0.008; J
3
=0.779 ± 0.010 (Наумов и др., 1987). l= –0.20 + 1.44h; h = 0.82 + 0.98b
(Наумов, Федяков, 1985). Максимальные размеры в Белом море: 8.4 × 5.7 × 4.5 мм (Наумов
и др., 1987).
Макушки прозогирные, смещены вперед.
Периостракум блестящий. Поверхность створок поделена на три поля: переднее, сред-
нее и заднее. Переднее поле равномерно покрыто радиальными ребрышками, среднее,
кроме линий, нарастания несет микроскопические
концентрические морщинки. Заднее по-
ле, отделенное от среднего четкой складкой, всегда орнаментировано ясной радиальной
исчерченностью.
Лунка и щиток неограниченны.
Лигамент наружный, погруженный.
Зубы замка отсутствуют. Края створок в области переднего и заднего нолей зазубрены.
Мантийная линия без синуса.
Отпечатки мускулов-замыкателей округлые, передний аддуктор значительно меньше
заднего и сильно сдвинут к переднему краю створок.
Палеонтологические находки. Известен с плейстоцена (Скарлато, 1981), в
отложениях Белого моря – с субатлантики (Говберг, 1968, 1970, 1973, 1975).
Распространение. Широко распространенный бореально-арктический циркум-
полярный вид. Встречен во всех северных морях России, море Бофорта, у берегов
Гренландии, Шпицбергена и Земли Франца-Иосифа. В Атлантическом океане рассе-
ляется на юг до мыса Хаттерас, в Тихом океане – до
зал. Петра Великого и зал.
Пьюджет-Саунд.
По материалам БентБоМ (встречен на 11 станциях), данным К. М. Дерюгина
(1928) (встречен на 8 станциях) и коллекций ЗИН РАН (7 пробы) в Белом море это –
относительно нечастый вид. Наибольшее количество находок приурочено к Двин-
скому заливу и прилегающим к нему участкам Бассейна. Обнаружен повсеместно,
кроме центральных
глубоководных частей Бассейна, Мезенского залива и Горла.
Впрочем, по данным К. М. Дерюгина (1928) встречен на обеих границах этого про-
лива (см. приложение 1, 9). Для более подробного обсуждения особенностей распро-
странения этого вида в Белом море данных недостаточно.
Основные экологические особенности. Относительно эвритопный, стено-
батный, предпочитающий илистые и песчаные грунты вид (Федяков
, 1986). По мате-
риалам БентБоМ отмечен на глубинах от 10 до 240 м при температуре от –1.5 до
10.0°С и солености от 20.2 до 28.8‰, в основном на илистых (50% находок) и сме-
шанных (40% находок) грунтах. Средняя биомасса этого вида на тех станциях, где он
обнаружен, составляет 0.303 ± 0.066г/м
2
, а средняя плотность поселения –
5.1 ± 1.0 экз./м
2
.
По материалам БентБоМ максимальная биомасса (0.748 г/м
2
при плотности посе-
ления 4 экз./м
2
) встречена на траверзе д. Кузомени на глубине 75 м на илистом грунте
MUSCULUS CORRUGATUS
205
при температуре –0.8°C в биоценозе Ophiacantha bidentata (табл. 38). Макси-
мальная плотность поселения (12 экз./м
2
при биомассе 0.356 г/м
2
) отмечена в Онеж-
ском заливе на глубине 10 м на илистом грунте в сообществе Yoldia hyperborea
(табл. 31).
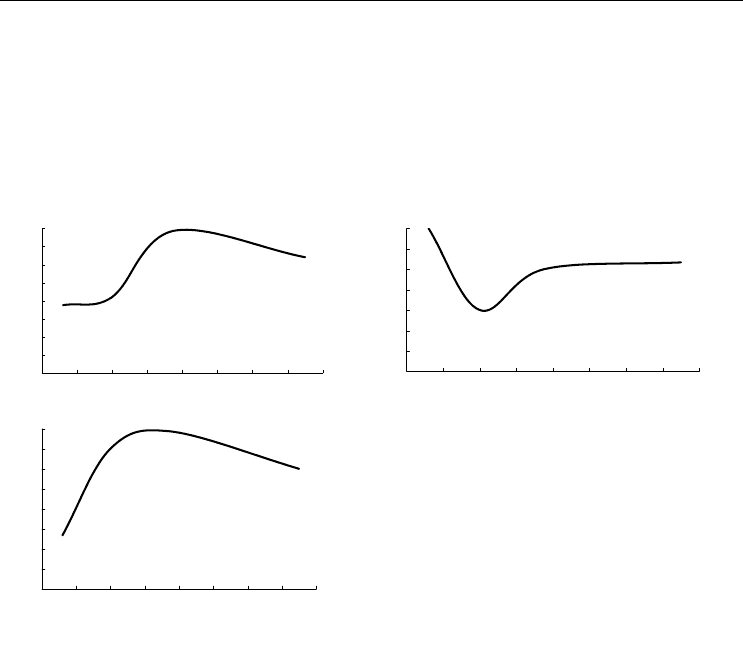
По материалам БентБоМ наибольшие значения биомассы наблюдаются на глуби-
не около 80 м, а плотность поселения максимальна на 20-метровой глубине и мини-
мальна приблизительно на 40 м. На этой последней глубине были обнаружены наи-
более крупные в среднем
экземпляры (рис. 103, А, Б, В).
Фильтрующий сестонофаг.
Зародышевая раковина имеет длину около 900 мкм (Наумов и др., 1987).
Продолжительность жизни в Белом море неизвестна.
А
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0 20 40 60 80 100 120 140 160
Б
0
1
2
3
4
5
6
7
0 20406080100120140160
В
0,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0 20 40 60 80 100 120 140 160
Рис. 103. Зависимость биомассы (А),
плотности поселения (Б) и
среднего веса
экземпляра (В)
Musculus corrugatus от
глубины.
Обозначения см. на рис. 92
Fig. 103. Biomass (А), density (Б) and
mean weight of a specimen (В) in
Musculus
corrugatu
s plotted against depth.
For legend see fig. 92

Глава 10. СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
206
Musculus laevigatus (Gray, 1824)
Modiola laevigata: Gray, 1824: 244
Modiolaria laevigata: Герценштейн, 1885: 644
Modiolaria discors (partim): Месяцев, 1931: 67-68; Горбунов, 1952: 232; Bernard, 1979: 27
Modiolaria discors laevigata: Дерюгин, 1928: 297; Горбунов, 1952: 232
Musculus (Musculus) laevigatus: Кафанов, 1991: 25
laevigatus: Dall, 1919: 5A; Филатова, 1948: 429, табл. 108, рис. 7; Ушаков, 1953: 262;
Скарлато, 1981: 229, рис. 127-129; Наумов и др., 1987: 223, рис. 133; Наумов, Федяков,
1994: 56; Galkin, Voronkov, 2001: 111
Диагноз (атлас, таблица II, 4).
Раковина вытянутая, светло-оливковая, или светло-коричневая, или зеленая, с
возрастом темнеет, равностворчатая, неравносторонняя, перламутровая.
J
1
= 0.633 + 0.008; J
3
= 0.651 ± 0.005 (Наумов и др., 1987). l = 0.40 + 1.52h;
h = 0.93 + 1.36b (Наумов, Федяков, 1985). Аллометрическая зависимость веса тела от
длины раковины W = 0.00027L
2.69
(Максимович, 1978б). Максимальные размеры в Белом
море: 35.9 × 28.5 × 12.0 мм (Наумов и др., 1987).
Макушки прозогирные, смещены вперед.
Периостракум блестящий. Поверхность створок поделена на три поля: переднее,
среднее и заднее. Переднее поле равномерно покрыто радиальными ребрышками;
среднее – гладкое; заднее, отделенное от среднего складкой, гладкое или покрыто тонкой
радиальной исчерченностью.
Лунка и
щиток не ограничены.
Лигамент наружный, погруженный.
Зубы замка отсутствуют. Края створок в области переднего и заднего полей зазубрены.
Мантийная линия без синуса.
Отпечаток заднего мускула-замыкателя округлый, переднего — вытянут вдоль края
раковины. Передний аддуктор значительно меньше заднего и сильно сдвинут к переднему
краю створок.
Ф. Бернард (Bernard, 1979), ссылаясь на работу А. С. Енсена (Jensen, 1912),
указавшего, что между Musculus discors и M. laevigatus имеются переходные формы,
сводит эти виды в синоним. Действительно, обе формы крайне сходны как по
конхиологическим признакам, так и по экологическим особенностям. Регрессионный
анализ пропорций раковины не выявляет различий между этими видами (см. главу 2),
однако по
габитуальным индексам они вполне достоверно различаются (см. диагнозы
обоих видов). На беломорском материале наблюдаются также устойчивые различия в
относительной величине переднего поля и в числе и размерах на них радиальных
ребрышек; кроме того, митилизация раковины M. laevigatus выражена ярче.
Названные конхиологические признаки значительно надежнее пропорций раковины,
по которым невозможно различить даже такие
виды, как Modiolus modiolus и Mytilus
edulis (см. главу 2), поэтому с мнением Ф. Бернарда нельзя согласиться.
Палеонтологические находки. Известен с плейстоцена (Скарлато, 1981), в
отложениях Белого моря – с суббореального времени (Невесский и др., 1977;
Говберг, 1973, 1975).
Распространение. Широко распространенный бореально-арктический циркум-
полярный вид. Встречен во всех северных морях России, море Бофорта, у берегов
Баффиновой Земли, Гренландии, Исландии, Шпицбергена и Земли Франца-Иосифа.
В Атлантическом океане расселяется на юг до Бискайского залива и штата Нью-
Йорк, в Тихом океане – до зал. Посьета и зал. Пьюджет-Саунд.
По материалам БентБоМ (встречен на 36 станциях), данным К. М. Дерюгина
(1928) (встречен на 2 станциях) и коллекций ЗИН РАН (39 проб)
в Белом море
распространен повсеместно за исключением Кандалакшского желоба (см.
приложение 1, 10). Единственная находка в этом районе (данные К. М. Дерюгина
1928), скорее всего, объясняется либо ошибкой в определении, что маловероятно,
либо сносом в этот район льдами или плавающими оторванными от субстрата
литоральными фукоидами. В Двинском заливе редок. В Онежском заливе по
данным
MUSCULUS LAEVIGATUS
207
Л. А. Кудерского (1961) встречен только в северной его части. Это неплохо
согласуется и с нашими данными.
Основные экологические особенности. Стенотермный умеренно
тепловодный, эврибатный, эвритопный вид, поселяющийся на любом виде субстрата
(Федяков, 1986). По материалам БентБоМ отмечен на глубинах от 2 до 70 м при
температуре от –0.9 до 11.8°С и солености от 18.2 до 27.9‰, в
основном на илистых
(27% находок) и смешанных (33% находок) грунтах. Средняя биомасса этого вида на
тех станциях, где он обнаружен, составляет 2.811 ± 1.457 г/м
2
, а средняя плотность
поселения – 27.7 ± 12.5 экз./м
2
.
По материалам БентБоМ максимальные биомасса (1.920 г/м
2
) и плотность
поселения (400 экз./м
2
) встречены у мыса Никодимского на глубине 5 м на
каменистом грунте при температуре 4.5°C солености 26.9‰ в биоценозе Phycodrys
(табл. 37).
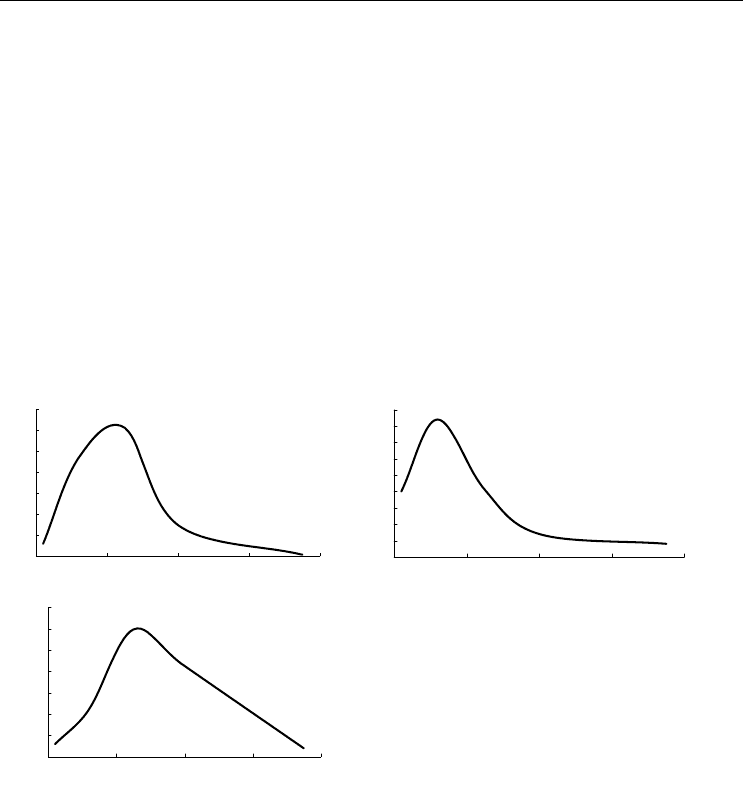
По материалам БентБоМ наибольшие значения биомассы наблюдаются на
глубине порядка 20–30 м, там же встречены и самые крупные в среднем экземпляры.
Максимальная плотность поселения обнаружена на глубине около 20 м (рис. 104, А,
Б, В).
Этот вид
служит первым промежуточным хозяином трематоды Prosorhynhcus
squamatus Odhner, 1905, приводящим к его паразитарной кастрации. Экстенсивность
заражения достигает 20%. Встречаются в нем и метацеркарии Gymnophallus delici-
osus (Зеликман, 1966).
Фильтрующий сестонофаг. Половой зрелости достигает на первом году жизни
(Максимович, 1980). Прикрепляется к талломам видов рода Laminaria, реже Fucus,
часто деформируя слоевище, биссусом, из нитей которого строит гнездо,
куда самки
откладывают кладки в виде белых закрученных шнуров (Матвеева, 1975, 1979).
Гнезда из биссуса строит на талломах. Иногда (но чаще, чем Musculus discors)
встречается на нитчатках и просто на грунте.
Нерест происходит летом. Диаметр зрелых яиц 300÷360 мкм (Кауфман, 1977) или
250 мкм (Матвеева, 1979). Слизистые кладки внутри гнезда встречаются в течение
всего года. Развитие без
пелагической личинки. Зародышевая раковина имеет длину
около 750 мкм.
Продолжительность жизни в Белом море – 2÷3 года (Максимович, 1980).
А
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 20406080
Б
0
5
10
15
20
25
30
35
40
45
0 20406080
В
0,000
0,005
0,010
0,015
0,020
0,025
0,030
0,035
0 20406080
Рис. 104. Зависимость биомассы (А),
плотности поселения (Б) и
среднего веса
экземпляра (В)
Musculus laevigatus от
глубины.
Обозначения см. на рис. 92
Fig. 104. Biomass (А), density (Б) and
mean weight of a specimen (В) in
Musculus
laevigatu
s plotted against depth.
For legend see fig. 92

Глава 10. СИСТЕМАТИЧЕСКОЕ ОПИСАНИЕ
208
Musculus niger (Gray, 1824)
Modiola nigra: Gray, 1824: 244
Modiolaria nigra: G. O. Sars, 1878: 31; Герценштейн, 1885: 645; Knipowitsch, 1902: 399;
Дерюгин, 1928: 298; Месяцев, 1931: 65; Горбунов, 1952: 231
Modiolaria nigra bullata: Дерюгин, 1928: 298, табл. 4, рис. 3, 4
Musculus discrepans: Филатова, 1948: 430, табл. 108, рис. 9
Musculus (Musculus) niger: Bernard, 1979: 27, fig. 37; Bernard, 1983: 20; Кафанов, 1991: 26
Musculus nigra: Ушаков, 1953: 263
Musculus niger: Скарлато, 1981: 231, рис. 131-133. Наумов и др., 1987: 224, рис. 134; Coan
et al., 2000: 169, pl. 25; Galkin, Voronkov, 2001: 111
Диагноз (атлас, таблица II, 5).
Раковина вытянутая, темно-оливковая до черной, равностворчатая, неравносторонняя,
перламутровая.
J
1
= 0.592 ± 0.007; J
3
= 0.627 ± 0.006 (Наумов и др., 1987). l = –1.99 + 1.94h;
h = 0.10 + 1.62b (Наумов, Федяков, 1985). Максимальные размеры в Белом море:
37.3 × 16.7 × 12.1 мм (Наумов и др., 1987).
Макушки прозогирные, смещены вперед больше, чем у других видов рода.
Периостракум блестящий. Поверхность створок поделена на три поля: переднее, сред-
нее и заднее. Переднее поле равномерно покрыто радиальными ребрышками
; среднее, по
крайней мере в области макушек, покрыто микроскопическими концентрическими мор-
щинками; заднее, неясно отделенное от среднего, несет тонкую (тоньше, чем у других ви-
дов рода) радиальную исчерченность, ослабевающую по направлению кпереди.
Лунка и щиток не ограничены.
Лигамент наружный, погруженный.
Зубы замка отсутствуют. Края створок в области переднего и заднего полей зазубрены.
Мантийная линия без синуса.
Отпечаток заднего аддуктора овальный, ориентирован вертикально, значительно
больше отпечатка переднего. Разница в размерах мускулов-замыкателей больше, чем у
других видов рода.
Палеонтологические находки. Известен с миоцена (Coan et al., 2000), в от-
ложениях Белого моря – с суббореального времени (Невесский и др., 1977; Говберг,
1968, 1970, 1973, 1975).
Распространение. Широко распространенный бореально-арктический цир-
кумполярный вид. Встречен во всех северных морях России, море Бофорта, у бере-
гов Гренландии, Исландии, Шпицбергена и Земли Франца-Иосифа. В Атлантиче-
ском океане расселяется на юг
до Северного моря и мыса Хаттерас, в Тихом океа-
не – до зал. Петра Великого и зал. Пьюджет-Саунд.
По материалам БентБоМ (встречен на 24 станциях), данным К. М. Дерюгина
(1928) (встречен на 23 станциях) и коллекций ЗИН РАН (30 пробы) в Белом море
обнаружен повсеместно, кроме кутовой части Онежского залива и северной части
Горла.
Возможно, это указывает на разобщенность поселений этого вида во внут-
ренних частях моря и в Мезенском заливе (см. приложение 1, 11).
Основные экологические особенности. Стенотермный умеренно тепло-
водный, относительно стенобатный, эвритопный вид, обитающий на илистых, пес-
чаных и каменистых грунтах (Федяков, 1986). По материалам БентБоМ отмечен на
глубинах от 6 до 180 м при температуре
от –0.1 до 13.0°С и солености от 23.0 до
28.0‰, в основном на илистых (38% находок) и смешанных (38% находок) грунтах.
Средняя биомасса этого вида на тех станциях, где он обнаружен, составляет
4.437 ± 1.477 г/м
2
, а средняя плотность поселения – 32.5 ± 17.5 экз./м
2
.
По материалам БентБоМ максимальная биомасса (16.560 г/м
2
при плотности по-
селения 24 экз./м
2
) обнаружена в Западной Соловецкой салме на глубине 20 м на
MUSCULUS NIGER
209
песчаном грунте при температуре 8.4°C в сообществе Modiolus modiolus
(табл. 36). Наибольшая плотность поселения (391 экз./м
2
) встречена в Кандалакш-
ском желобе на глубине 180 м на илистом грунте при температуре около 0°C соле-
ности около 28‰ в биоценозе Portlandia arctica (табл. 39).
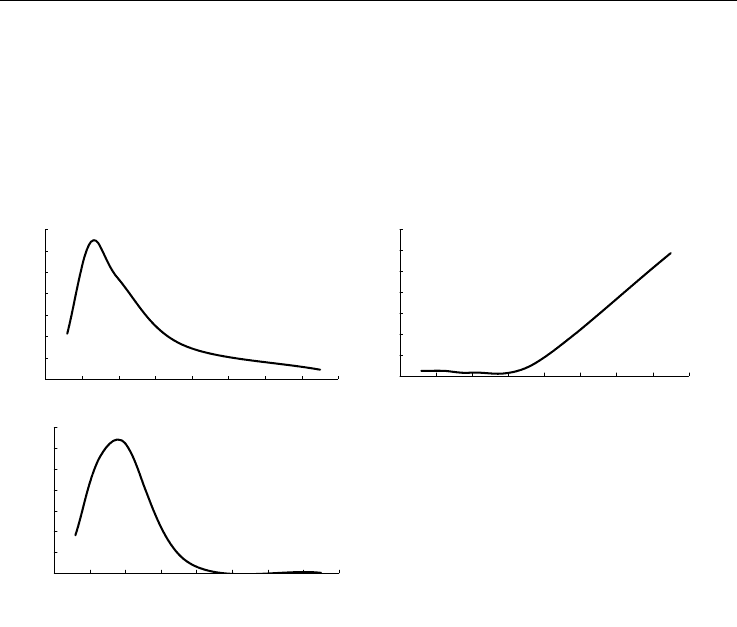
По материалам БентБоМ наибольшие значения биомассы и наиболее крупные в
среднем экземпляры встречены на глубинах около 40 м, в то время как плотность
поселения этого вида крайне низка до глубины приблизительно 60
м, а с дальней-
шим увеличением глубины начинает быстро расти (рис. 105, А, Б, В).
Фильтрующий сестонофаг. Строит гнезда из биссуса и алевро-пелитовых частиц
грунта.
Сроки и способ размножения неизвестны. Зародышевая раковина имеет длину
около 900 мкм (Наумов и др., 1987).
Продолжительность жизни в Белом море неизвестна.
А
0
1
2
3
4
5
6
7
0 20406080100120140160
Б
0
50
100
150
200
250
300
350
0 20 40 60 80 100 120 140 160
В
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 20 40 60 80 100 120 140 160
Рис. 105. Зависимость биомассы (А), плот-
ности поселения (Б) и
среднего веса эк-
земпляра (В)
Musculus niger от глубины.
Обозначения см. на рис. 92
Fig. 105. Biomass (А), density (Б) and mean
weight of a specimen (В) in
Musculus niger
plotted against depth.
For legend see fig. 92