Murray J.D. Mathematical Biology: I. An Introduction
Подождите немного. Документ загружается.


416 11. Reaction Diffusion, Chemotaxis, Nonlocal Mechanisms
Section 11.4. If we now include a reaction or dynamics term f (n) in (11.59) we get the
generalised reaction diffusion equation equivalent to (11.14). With the one space dimen-
sional scalar version of (11.59) and a logistic growth form for f (n), Cohen and Murray
(1981) have shown that the equation can exhibit steady state spatially inhomogeneous
solutions. Lara-Ochoa (1984) analysed their model in a two-dimensional setting and
showed that it reflects certain morphogenetic aspects of multicellular systems formed
by motile cells.
Exercises
1 Let p(x, t) be the probability that an organism initially at x = 0isatx after a
time t. In a random walk there is a slight bias to the right; that is, the probabilities of
moving to the right and left, α and β, are such that α −β = ε>0, where 0 <ε 1.
Show that the diffusion equation for the concentration c(x, t) = Qp(x, t),whereQ
particles are released at the origin at t = 0, is
∂c
∂t
+ V
∂c
∂x
= D
∂
2
c
∂x
2
,
where V and D are constants which you should define.
2 In a one-dimensional domain suppose insects are attracted to the origin x = 0andare
convected there by a constant velocity V . If the population pressure is approximated
by a density-dependent diffusion coefficient D(n) = D
0
(n/n
0
)
m
,wheren is the
population density and D
0
, n
0
and m are positive constants, show that the model
equation for dispersal, in the absence of any population growth, is
∂n
∂t
=−
∂ J
∂x
=
∂
∂x
[V sgn(x)n]+D
0
∂
∂x
n
n
0
m
∂n
∂x
.
Show that if n → 0, ∂n/∂x → 0as|x |→∞a spatially inhomogeneous steady
state population density exists and can be represented by
n(x) = n
0
1 −
mV|x |
D
0
1/m
, if |x |≤
D
0
mV
= 0, if |x | >
D
0
mV
.
3 The larvae of the parasitic worm (Trichostrongylus retortaeformis) hatch from eggs
in sheep and rabbit excreta. The larvae disperse randomly on the grass and are conse-
quently eaten by sheep and rabbits. In the intestines the cycle starts again. Consider
the one-dimensional problem in which the larvae disperse with constant diffusion
and have a mortality proportional to the population. Show that n satisfies
∂n
∂t
= D
∂
2
n
∂x
2
−µn, D > 0,µ>0,
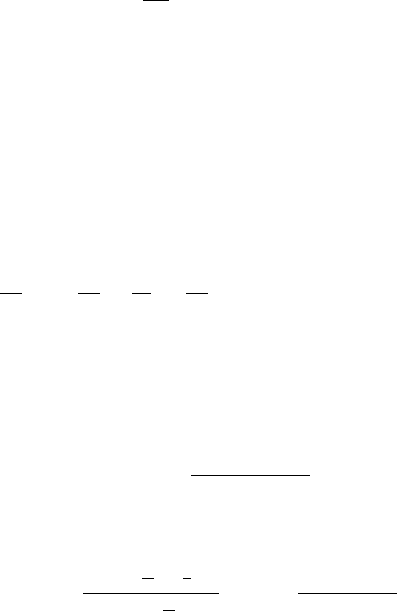
Exercises 417
where n is the larvae population. Find the population distribution at any x and t
arising from N
0
larvae being released at x = 0att = 0. Show that as t →∞the
population dies out.
If the larvae lay eggs at a rate proportional to the population of the larvae,
that is,
∂ E
∂t
= λn,λ>0,
where E(x, t) is the egg population density, show that in the limit as t →∞a
nonzero spatial distribution of eggs persists. [The result for E(x, t) is an integral
from which the asymptotic approximation can be found using Laplace’s method
(see, for example, Murray’s 1984 Asymptotic Analysis): the result gives E(x, t) ∼
O(exp[−(µ/D)
1/2
|x |]) as t →∞.]
4 Consider the density-dependent diffusion model for insect dispersal which includes
a linear death process which results in the following equation for the population
n(x, t),
∂n
∂t
= D
0
∂
∂x
n
n
0
m
∂n
∂x
−µn, D
0
> 0,µ>0.
If Q insects are released at x = 0att = 0, that is, n(x, 0) = Qδ(x), show, using
appropriate transformations in n and t, that the equation can be reduced to an equiva-
lent equation with µ = 0. Hence show that the population wavefront reaches a finite
distance x
max
from x = 0ast →∞,where
x
max
=
r
0
(µmτ
0
)
1/(m+2)
,
where
r
0
=
Q(
1
m
+
3
2
)
π
1/2
n
0
(
1
m
+1)
,τ
0
=
r
2
0
m
2D
0
(m +2)
,
where is the Gamma function.
12. Oscillator-Generated Wave Phenomena and
Central Pattern Generators
In Chapter 11 we saw how diffusion, chemotaxis and convection mechanisms could
generate spatial patterns; in Volume II we discuss mechanisms of biological pattern for-
mation extensively. In Chapter 13, and Chapter 1 and Chapter 13, Volume II we show
how diffusion effects, for example, can also generate travelling waves, which have been
used to model the spread of pest outbreaks, travelling waves of chemical concentration,
colonization of space by a population, spatial spread of epidemics and so on. The exis-
tence of such travelling waves is usually a consequence of the coupling of various effects
such as diffusion or chemotaxis or convection. There are, however, other wave phenom-
ena of a quite different kind, called kinematic waves, which exhibit wavelike spatial
patterns, which depend on the coupling of biological oscillators whose properties relat-
ing to phase or period vary spatially. The two phenomena described in this chapter are
striking, and the models we discuss are based on the experiments or biological phenom-
ena which so dramatically exhibit them. The first involves the Belousov–Zhabotinskii
reaction and the second, which is specifically associated with the swimming of, for ex-
ample, lamprey and dogfish, illustrates the very important concept of a central pattern
generator. The results we derive here apply to spatially distributed oscillators in general.
12.1 Kinematic Waves in the Belousov–Zhabotinskii Reaction
When the reactants in the oscillating Belousov reaction involve an iron catalyst (with
Fe
2+
going to Fe
3+
and vice versa) the oscillations are dramatically illustrated with an
appropriate dye which reflects the state of the catalyst: the colour change is from red
(or rather a reddish orange) to blue. When the reactants are left unstirred in a vertical
cylindrical tube horizontal bands of blue and red form. These bands usually start to
appear at the bottom of the cylinder and move slowly upwards with successive bands
moving progressively more slowly. Eventually the cylinder is filled by these bands but
with a nonuniform density, the closer to the bottom the denser the wave packing. Dif-
fusion plays a negligible role in the formation and propagation of these bands, unlike
the waves we discuss later. Beck and V
´
aradi (1972) provided a kinematical explanation
for these spatial patterns of bands. The analysis explaining them, which we give here,
is that of Kopell and Howard (1973). Although the analysis was originally given for the
bands observed in the Belousov reaction, and the experimental results shown in Fig-
ure 12.1(b) are also for this reaction, the phenomenon and analysis apply equally to any
12.1 Belousov–Zhabotinskii Reaction Kinematic Waves 419
biological oscillator under similar circumstances. The important point to note is that
spatial patterns can be obtained without diffusion, convection or chemotaxis playing
any role.
Consider each position in the vertical cylinder to be an independent oscillator with
period T , which may be a function of position. If these independent oscillators are
out of phase or have different frequencies then spatial patterns will appear simply as a
consequence of the spatial variation in the phase or frequency. (A simple but illustrative
physical demonstration of the phenomenon is given by a row of simple pendula all
hanging from the same horizontal rod but with a very slight gradient in their lengths.
The slight gradient in their lengths gives a slight gradient in their periods. If they are all
set swinging at the same time, then after a very short time it looks as if there is a wave
propagating along the line of pendula, the wavelength of which gets smaller and smaller
with time.)
Returning to the Belousov oscillator, the cause of a gradient in phase or frequency
can be due to a concentration gradient in one of the chemicals, or a temperature vari-
ation. The experiments (see Figure 12.1(b)) carried out by Kopell and Howard (1973)
used the former while Thoenes (1973) used the latter. The vertical chemical concentra-
tion gradient was in sulphuric acid. This resulted in a monotonic gradient in the period
of oscillation and horizontal bands appeared quite quickly, moved slowly upwards and
after a few minutes filled the cylinder. It is clear that if a barrier, impermeable to any
of the chemicals, were put in the cylinder it would neither affect the pseudowave prop-
agation nor the density of bands: spatial transport processes are simply not involved in
the generation of this spatial pattern. These ‘waves’ are indeed only pseudowaves since
nothing is actually being transported.
Let z be the spatial coordinate measured vertically from the bottom of the cylinder,
takentobez = 0, and the cylinder height to be normalised so that the top is z = 1.
Because of the initial concentration gradient there is a gradient in the oscillator period.
At position z let the period of oscillation be T (z) defined for all 0 ≤ z ≤ 1. We char-
acterise the state of the oscillator by a 2π-periodic function of its phase denoted by
φ(z, t). In the Belousov reaction, for example, the front of the wave, defined as the
point where φ has a specific value, can be distinguished by the sharp blue front. Let the
initial distribution of the phases be φ
0
(z). We can then represent the phase φ(z, t) by
φ(z, t) = ψ(z, t) +φ
0
(z), ψ(z, 0) = 0, (12.1)
where ψ(z, t) is a function which increases by 2π if the time t increases by the periodic
time T (z);thatis,
φ(z, t + T (z)) = 2π +ψ(z, t) +φ
0
(z).
Let us now take some reference phase point say, φ = 0. Define t
∗
(z) as the time at
position z at which the phase is zero; that is, it satisfies
0 = φ(z, t
∗
(z)) = ψ(z, t
∗
(z)) +φ
0
(z). (12.2)
Then, for any integer n and time t = t
∗
(z) + nT(z) we have

420 12. Oscillator-Generated Wave Phenomena
φ(z, t
∗
+nT) = ψ(z, t
∗
+nT) + φ
0
(z)
= 2nπ +ψ(z, t
∗
) +φ
0
(z)
= 2nπ,
(12.3)
using (12.1) and (12.2). So, in the (z, t) plane the point (z, t) which corresponds to the
phase 2nπ moves on the curve given by
t = t
∗
(z) + nT(z). (12.4)
We can continue the analysis with complete generality but it is just as instructive
and easier to see what is going on if we are more specific and choose T (z) tobe,say,a
smooth monotonic increasing function of z in 0 ≤ z ≤ 1. Also for simplicity, let us take
the initial distribution of phases to be a constant, which we can take to be zero; that is,
φ
0
(z) = 0. This means that at t = 0 all the oscillators are in phase. From the definition
of t
∗
(z) in (12.2) this means that t
∗
(z) = 0.
If we now define
t
n
(z) = nT(z) (12.5)
then (12.3) gives
φ(z, t
n
(z)) = 2nπ. (12.6)
This means that t
n
(z) is the time at which the nth wavefront passes the point z in the
cylinder. The velocity v
n
(z) of this nth wavefront is given by the rate of change of the
position of the front. That is, using the last two equations,
v
n
(z) =
dz
dt
φ=2nπ
=
dt
n
(z)
dz
−1
=
1
nT
(z)
. (12.7)
With T (z) a monotonic increasing function of z, T
(z)>0andsothenth wave-
front, the leading edge say, starts at z = 0 at time t = nT(0) and, from (12.7), propa-
gates up the cylinder at 1/n times the velocity of the first wave. This nth wave reaches
z = 1, the top of the cylinder, at time t = nT(1). Since the T (z) we have taken here is
a monotonic increasing function of z there will be more and more waves in 0 ≤ z ≤ 1
as time goes on, since, with the velocity decreasing proportionally to 1/n from (12.7),
more waves enter at z = 0thanleavez = 1 in the same time interval.
Let us consider a specific example and take φ
0
(z) = 0andT (z) = 1 + z.Sothe
phase 2nπ moves, in the (z, t) plane, on the lines t = n(1 +z), and the phase φ(z, t) =
2πt/T (z) = 2πt/(1 + z). From (12.7) the velocity of the nth wave is v
n
= 1/n.The
space-time picture of the wavefronts, given by φ = 2nπ, is illustrated in Figure 12.1(a).
From the figure we see that at time t = 1thewaveφ = 2π enters the cylinder at z = 0
and moves up with a velocity v
1
= 1. At t = 2 the wave with phase 4π enters the
cylinder at z = 0: it moves with velocity v
2
= 1/2. At t = 3 the wave with φ = 6π
moves with velocity v
3
= 1/3 and so on. The first wave takes a time t = 1 to traverse
the cylinder, the second takes a time t = 2 and so on. It is clear that as time goes on
12.1 Belousov–Zhabotinskii Reaction Kinematic Waves 421
Figure 12.1. (a) Wavefronts for the period distribution T (z) = 1+z and initial phase distribution φ
0
(z) = 0.
Note how many waves there are for different t’s: for t = 1.5 there is 1 wave in the cylinder, while for t = 7.5
there are 4 waves. (b) The experimental space–time situation equivalent to the theoretical results in (a). The
experiments were carried out for a Belousov reaction with an initial sulphuric acid gradient in the cylinder.
The total time in the figure is approximately 7 minutes and the vertical height about 20 cm; the figure is a
sketch from a negative. (After Kopell and Howard 1973)
more and more waves are in 0 ≤ z ≤ 1. This is clear from Figure 12.1(a) and the
experimental counterpart given in Figure 12.1(b): for example, at t = 3.5therearetwo
waves while at t = 7.5 there are 4. From the figure, it is also clear that as time increases
the waves are progressively more tightly packed nearer the bottom, z = 0.
Suppose the initial phases φ
0
(z) = 0; then, t
∗
(z) = 0 and from (12.4), T (z) =
[t − t
∗
(z)]/n. So, asymptotically for large time t, T (z) ∼ t/n, and the above analysis
for the illustrative example in which φ
0
(z) = 0 still applies asymptotically in time. Since
it is unlikely that the experimental arrangement which gave rise to Figure 12.1(b) had
a strictly linear T (z), such as used in the analysis for Figure 12.1(a), the experimental
results illustrate this asymptotic result quite dramatically.
The biological implications of the above analysis are of considerable importance.
Since biological oscillators and biological clocks are common, a time-varying spatial
422 12. Oscillator-Generated Wave Phenomena
pattern may be a consequence of a spatial variation in the oscillator parameters and not,
as might be supposed, a consequence of some reaction diffusion situation or other such
pattern formation mechanism such as we shall consider in detail in later chapters.
In this section the wave pattern is a continuously changing one. In the following
sections we investigate the possibility of a more coherent wave pattern generator of
considerable importance.
12.2 Central Pattern Generator: Experimental Facts in the
Swimming of Fish
A fish propels itself through water by a sequence of travelling waves which progress
down the fish’s body from head to tail and its speed is a function of the wave frequency.
It is the network of neurons arrayed down the back that controls the muscle movements
which generate the actual waves and coordinate them to produce the right effect. It is
a widely held lay belief that in mammals the generator, or rather the controlling nerve
centre for the rhythmic control of these waves, is the brain. However, in many animals
swimming occurs after the spinal cord has been severed from the brain—the technical
term is spinal transection. In the case of the dogfish, for example, the phenomenon has
been known since the end of the 19th century. The swimming movement observed in
such situations shows the proper intersegmental muscle coordination.
The basis for the required rhythmic behaviour and its intersegmental coordination is
a central network of neurons in the spinal cord. It is known that there are neural networks
which can generate temporal sequences of signals, which here produce the required
cyclic patterns of muscle activity. Such networks are called central pattern generators
and by definition require no external input control for them to produce the required
rhythmic output. It is obvious how important it is to understand such neural control
of locomotion. However to do so requires modelling realism and, at the very least,
detailed information from experiments. The recent book on neural control of rhythmic
movements, edited by Cohen et al. (1988), is specifically about the subject matter of this
section and the theory and modelling chapters by Kopell (1988) and Rand et al. (1988)
are particularly relevant.
In the case of higher vertebrates there are possibly millions of neurons involved. So
it is clear that experimentation and its associated modelling should at least start with as
small a spinal cord as possible but one which still exhibits this posttransection activity.
Such neural activity, which produces essentially normal swimming, is called ‘fictive
swimming’ or ‘fictive locomotion.’ This description also includes the situation where,
even when the muscles which produce the actual locomotion are removed, the neural
output from the spinal cord is the same as that of an intact swimming fish.
Grillner (1974), Grillner and Kashin (1976) and Grillner and Wall
´
en (1982) present
good experimental data on the dogfish. Kopell (1988) uses this work as a case study for
the theory described in detail in her paper. The lamprey, which is rather a primitive
vertebrate, was the animal used in a series of interesting and illuminating experiments
by Cohen and Wall
´
en (1980) (see also Cohen and Harris-Warrick 1984 and references
given there). They studied a specific species of the lamprey which varies from about
13–30 cm in length and has a spinal cord about 0.3 mm thick and 1.5 mm wide. Its
12.2 Central Pattern Generator 423
Figure 12.2. (a) Schematic diagram of the exposed lamprey spinal cord and the experimental arrangement.
The preparation was pinned through membrane tissue as shown in the schematic cross-section in (b). There
are ventral roots (VR) at each segment and the electrical activity of these was measured by the electrodes.
(From Cohen and Wall
´
en 1980)
advantages (or disadvantages from the lamprey’s point of view) are that it has relatively
few cells, but still has the necessary basic vertebrate organisation, and it exhibits fictive
swimming behaviour. In the experiments of Cohen and Wall
´
en (1980) and Cohen and
Harris-Warrick (1984), dissected lengths of spinal cord from about 25–50 segments
were used: the lamprey has about 100. The animal was decapitated and the spinal cord
exposed but with most of the musculature intact. Motoneuron activity was monitored
with electrodes placed on two opposing ventral roots of a single segment; Figure 12.2
shows schematically the experimental setup of the spinal cord.
In these experiments the cord was placed in a saline solution and the fictive swim-
ming, that is, the periodic rhythmic activity, was induced chemically (by L-DOPA or by
the amino acid D-glutamate); the fictive swimming can go on for hours. The ventral root
(VR) recordings from the electrodes showed alternating bursts of impulses between the
left and right VR of a single segment. That is, the periodic bursts on either side of a
segment are 180
◦
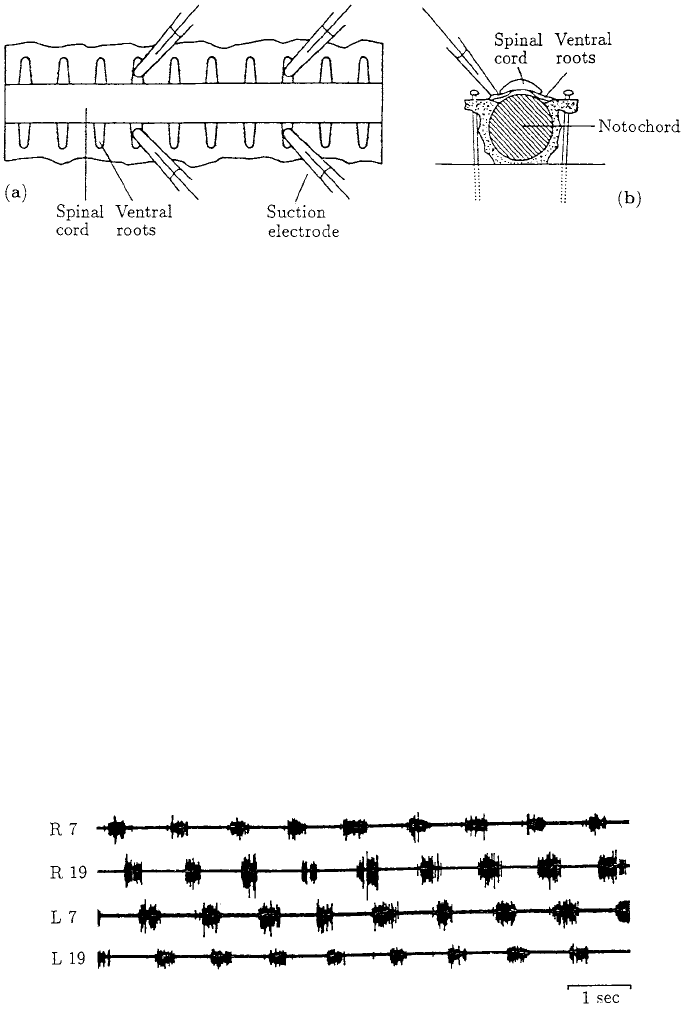
out of phase. Figure 12.3 illustrates the VR activity obtained from the
left and right sides of the ventral roots from two different segment levels in the cord. An
Figure 12.3. Bursting activity recorded from the left (L) and right (R) sides of the ventral roots (VR) at
segments 7 and 19 as measured from the head-end of the specimen, which consisted of 27 segments. (From
Cohen and Wall
´
en 1980) The time between bursts is approximately 1 sec. Note the approximately constant
phase lag as you go from segment 7 to 19.

424 12. Oscillator-Generated Wave Phenomena
Figure 12.4. Typical swimming pattern illustrating
a propagating wave generated by a ventral root
output such as illustrated in Figure 12.3.
important point to note, and which we use in the model, is that the left and right VR of
a segment are like individual oscillators which are phase locked 180
◦
out of phase. This
intrasegmental coupling is very stable.
The results in Figure 12.3 are from an isolated piece of spinal cord consisting of
27 segments and with the numbering starting from the head side; the recordings were
taken at segments 7 and 19. The period of the bursts of activity was about 1 per second.
Another point to note is the nearly constant phase lag between the two segments: the lag
between the right VR of segment 7 and segment 19 is to a first approximation the same
as between the left VR of segments 7 and 19. In Section 12.4 we incorporate this type of
behaviour in one of the specific cases in the model developed below. A piece of spinal
cord of only about 10 segments can produce a stable neural fictive swimming output.
This periodic activity of the isolated lamprey spinal cord, which is directly related
to the undulatory wavelike movements of the swimming fish, schematically shown in
Figure 12.4, is the phenomenon controlled by the central pattern generator, which we
now wish to model. Various models can be suggested for the generation of these pat-
terns; see, for example, Kopell (1988). In the following Sections 12.3 and 12.4 we de-
scribe in detail a model proposed by Cohen et al. (1982); it has been used with consid-
erable success to explain certain experimental results associated with selective surgical
lesions in the spinal cord (Cohen and Harris-Warrick 1984).
12.3 Mathematical Model for the Central Pattern Generator
The basic characteristics of the phenomenon, as exemplified by the experiments and
indicated in particular by Figure 12.3, are that the left and right VR of a segment are
phase locked oscillators and that there is approximately a constant phase lag from the
head to tail of the spinal cord. The key assumptions in the model are: (i) Each segment
in the back has associated with it a pair of neuronal oscillators each of which exhibits,
in isolation, a stable limit cycle periodic oscillation. The amplitude of the oscillation
depends only on internal parameters, and is not usually affected by external factors
such as drugs or electrical stimulations. (ii) Each of the oscillators is coupled to its
nearest neighbour but with the possibility of long range coupling; there is experimental
evidence for the latter in Buchanan and Cohen (1982).

12.3 Mathematical Model for the Central Pattern Generator 425
We saw in Chapter 7 on biological oscillators that many biochemical reaction sys-
tems can exhibit stable limit cycle periodic oscillations. It is such a biological oscillator,
or one coupled to some neuronal electrical property, which we envisage to be the driving
force in each of the oscillators associated with the spinal segments. It is not necessary
to know the actual details of the biological oscillator for our model here—we do not
know what it is in fact. As a preliminary to studying the intersegmental linking of the
oscillators we first consider a single oscillator to set up the mathematical treatment and
notation and introduce the analytical procedure we use.
Single Oscillator and Oscillator Pair
Denote the vector of limit cycle variables of relevance by the vector
x(t) = (x
1
(t), x
2
(t),... ,x
n
(t)). (12.8)
Again we do not know what quantity it is that oscillates, only that something does
which gives rise to the periodic VR neuronal activity which is observed experimentally.
For example, x(t) could include the level of the neurotransmitter substance and the peri-
odically varying electric potential. We denote the vector differential equation governing
the limit cycles by
dx
dt
= f(x), (12.9)
where t denotes time. Just as we do not need to know the specific biological oscillator
involved, we do not require the detailed functional form of the function f(x).Letus
consider the limit cycle to be the closed orbit γ in the phase space. Using this curve
as the local coordinate system we can think of the periodic limit cycle as having a
phase θ which goes from 0 to 2π as we make a complete circuit round the closed
orbit γ . Assume that at some point P on the closed curve the bursting, which is observed
experimentally by the electrodes, occurs at the phase θ = 0. Starting at P, the phase
increases from 0 and reaches 2π when we get back to P, where bursting again occurs.
Let us further assume that the coordinate system for the limit cycle is chosen so that the
speed of the solution round γ , as measured now by dθ/dt, is constant.
The above idea of representing a limit cycle in terms of the phase can be illus-
trated by the following, admittedly contrived, simple pedagogical example. Consider
the differential equation system given by
dx
1
dt
= x
1
(1 − ρ) − ωx
2
,
dx
2
dt
= x
2
(1 − ρ) +ωx
1
,
ρ = (x
2
1
+ x
2
2
)
1/2
,
(12.10)
where ω is a positive constant. Although the solution of this system can be obtained triv-
ially, as we see below, a formal phase plane analysis (see Appendix A) of (12.10) shows
that (0, 0) is the only singular point and it is an unstable spiral, spiralling anticlockwise.
A confined set can be found (just take ρ large), so by the Poincar
´
e–Bendixson theorem
a limit cycle periodic solution exists and is represented by a closed orbit in the (x
1
, x
2
)