Лекция- Обмен нуклеотидов
Подождите немного. Документ загружается.

ЛЕКЦИЯ
АНАБОЛИЗМ И КАТАБОЛИЗМ ПУРИНОВЫХ И
ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Переваривание нуклеиновых кислот начинается в желудке, где под
действием желудочного сока происходит отщепление белковых компонентов
от нуклеопротеинов (рис.1.).
ДНК-азы и РНК-азы панкриотического сока, являясь эндонуклеазами,
гидролизуют макромолекулы до олигонуклеотидов. Последние, под
действием фосфоэстераз, расщепляются до смеси 3’- и 5’-мононуклеотидов.
Нуклеотидазы и неспецифические фосфатазы гидролитически отщепляют
фосфатные остатки нуклеотидов, превращая их в нуклеозиды. Нуклеозиды
либо всасываются клетками тонкого кишечника, либо расщепляются
нуклеозидфосфорилазами кишечника с образованием рибозо- или
дезоксирибозо-1-фосфатов, пуриновых и пиримидиновых оснований.
Как правило, образовавшиеся пурины и пиримидины очень мало
используются для синтеза нуклеиновых кислот и в энтероцитах под
действием активной ксантиноксидазы превращяют пуриновые основания в
мочевую кислоту, которая в дальнейшем выводятся из организма с мочой.
Пиримидины под действием микрофлоры кишечника расщепляются до NН
3
,
СО
2
, β-аланина и β-аминоизобутирата.
Различные клетки организма синтезируют до 90% пуриновых и
пиримидиновых нуклеотидов de novo.
Синтез пуриновых нуклеотидов de novo
Было установлено, что в формировании пуринового кольца принимают
участие аминокислоты: аспарагиновая, глутаминовая и глицин, СО
2
и два
одноуглеродных производных тетрагидрофолата (N
1
-аспартат, С
2
–N
10
-
формил-Н
4
-фолат, N
3
и N
9
-амидный азот глутамина, N
7
, С
4
и С
5
–глицин, С6-
СО
2
, С
8
-N
5
,N
10
-метенил-Н
4
-фолат).
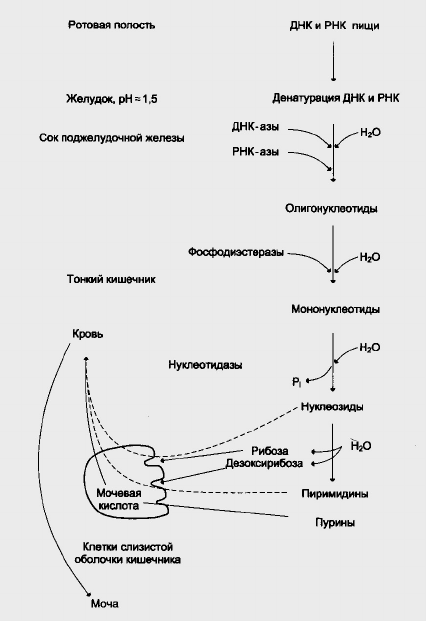
Рис. 1. Переваривание нуклеиновых кислот в пище.
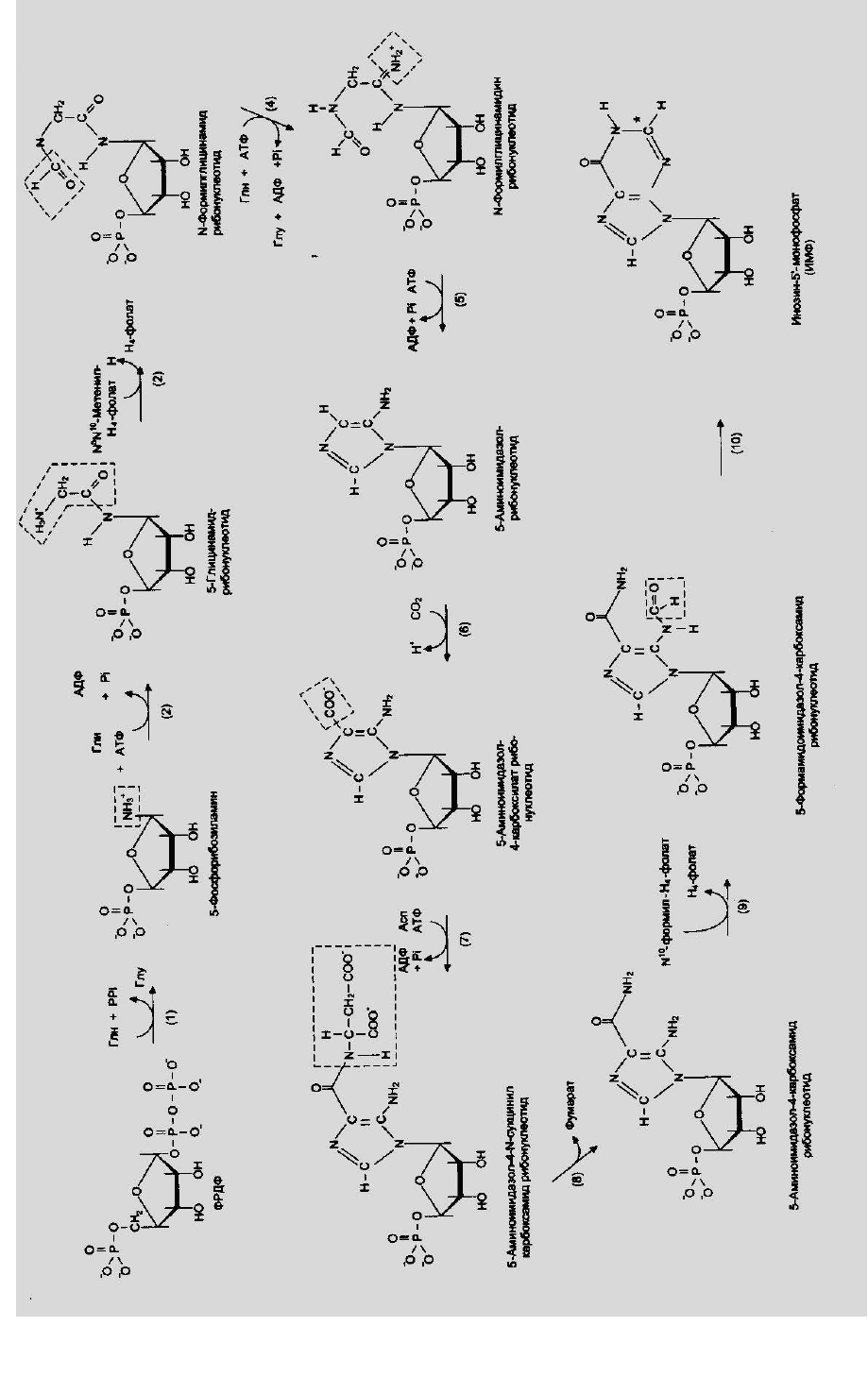
Рис.2. Синтез пуриновых нуклеотидов de novo.
Общим предшественником синтеза всех пуриновых нуклеотидов
является инозин-5-монофосфат (ИМФ), содержащий в качестве азотистого
основания гипоксантин.
Первая реакция синтеза ИМФ требует наличия ФРПФ-
фосфорибозилтрифосфата, который занимает центральное место не только в
анаболизме пуринов, но необходим при синтезе пиримидинов (рис.2.).
Он образуется за счет переноса β,γ-пирофосфатного остатка АТФ на
рибозил-5-фосфат, в реакции катализируемой ФРПФ-синтетазой.
Источником рибозил-5-фосфата может быть пентозофосфатный путь
превращения глюкозы или катаболизма нуклеотидов, в ходе которого под
действием нуклеозидфосфорилазы первоначально образуется рибозо-1-
фосфат, а затем под действием соответствующей мутазы фосфорный остаток
переносит в 5-положение.
Анаболический путь синтеза ИМФ заканчивается образованием
гипоксантина. Этот путь значительно более сложный, чем путь синтеза
пиримидиновых оснований.
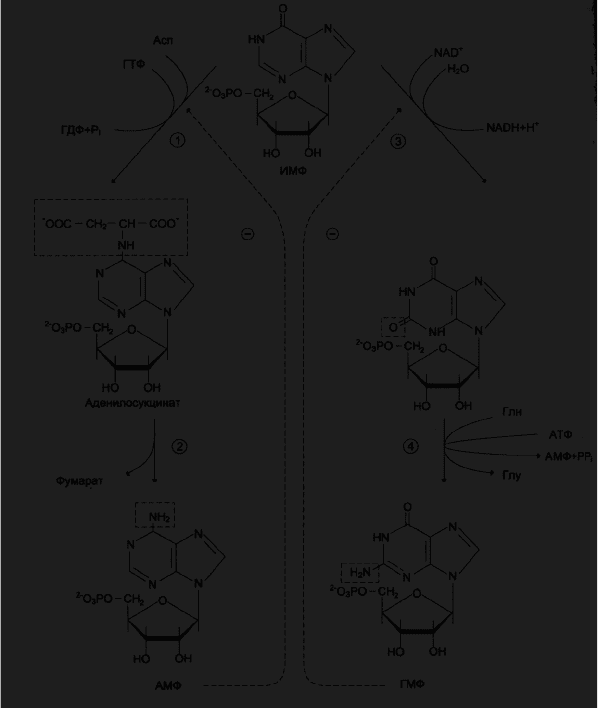
Рис.3.Синтез АМФ и ГМФ из ИМФ. 1- аденилосукцинатсинтетаза; 2-
аденилосукциназа; 3- ИМФ-дегидрогеназа, 4 –ГМФ-синтетаза.
Синтез пуриновых оснований через образование ИМФ требует затраты
значительного количества энергии в форме АТФ. Так, на синтез циклической
структуры пуринов затрачивается 5 молекул АТФ на каждую молекулу АМФ
или ГМФ. ИМФ в основном используется для синтеза АМФ или ГМФ.
Небольшое количество этого продукта обнаруживается также в т-РНК в
качестве одного из минорных оснований (рис.3.).
Известен другой путь образования пула пуриновых оснований – путь
реутилизации пуриновых оснований, образовавшихся в процессе распада
эндогенных или экзогенных нуклеотидов. Очевидно, эти реакции можно
рассматривать как «сберегающие», использующие пуриновые кольца до их
превращения в ксантин и затем в мочевую кислоту перед экскрецией.
Регуляция синтеза пуриновых оснований связана с концентрацией
ФРПФ, которая, в свою очередь, зависит от скорости его синтеза, утилизации
и разрушения (рис.4.).
Количество ФРПФ определяется наличием рибозо-5-фосфата и
активностью ФРПФ-синтетазы - фермента чувствительного к концентрации
фосфатов и пуриновых нуклеотидов.
Внутриклеточная концентрация ФРПФ строго регулируется и обычно
мала. ФРПФ-синтетаза является аллостерическим ферментом и активируется
неорганическим фосфором и ингибируется пуриновыми нуклеозид-моно-,
ди- и трифосфатами, при этом эффективность ингибирования распределяется
в следующем порядке: НМФ>НДФ>НТФ.
ФРПФ служит не только субстратом, но и аллостерическим
активатором второй реакции синтеза пуринонуклеотидов de novo, которую
катализирует амидофосфорибозилтрансфераза.
Пуриновые нуклеотиды, особенно АМФ и ГМФ, по механизму
отрицательной обратной связи ингибируют амидофосфорибозилтрансферазу,
которая катализирует первую специфическую реакцию синтеза пуриновых
оснований de novo.
Метаболическая цепь образования АМФ и ГМФ de novo регулируется в
месте её разветвления: АМФ ингибирует аденилосукцинатсинтетазу, а ГМФ-
реакцию образования ксантиловой кислоты, которую катализирует
ИМФ дегидрогеназа.
Перекрёстная регуляция путей использования ИМФ служит для
того,чтобы снизить синтез одного пуринового нуклеотида при дефиците
другого.
Помимо ферментов основного пути синтеза пуриновых нуклеотидов de
novo, регулируется также активность ферментов «запасных» путей:
аденинфосфорибозилтрансфераза ингибируется АМФ, а гипоксантин-
гуанинрибозилтрансфераза – ИМФ и ГМФ.
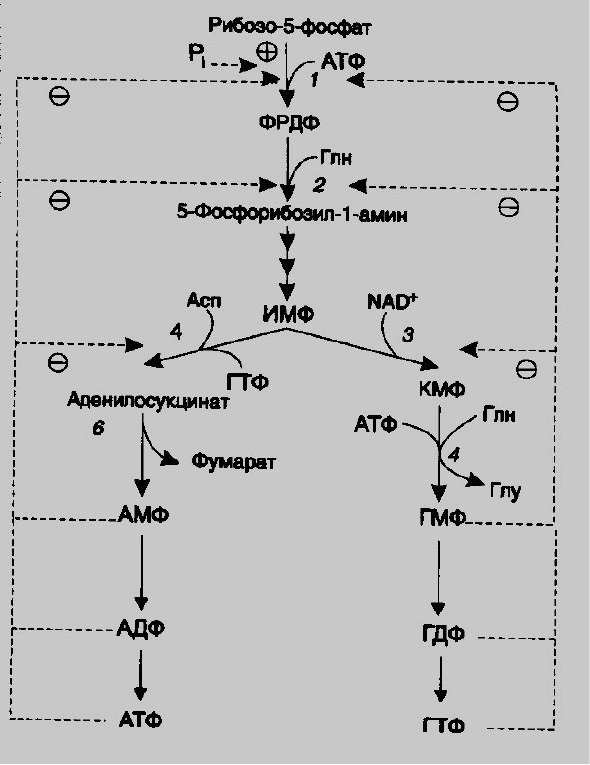
Рис.4. Регуляция синтеза пуриновых нуклеотидов. 1- ФРПВ-синтетаза; 2 –
амидофосфорибозилтрансфераза; 3 – ИМФ-дегидрогеназа; 4-
аденилосукцинатсинтетаза.
Катаболизм пуриновых нуклеотидов
У человека основным продуктом катаболизма пуриновых нуклеотидов
является мочевая кислота (рис.5.). Её образование идет путем
гидролитического отщепления фосфатного остатка от нуклеотидов с
помощью нуклеотидаз или фосфатаз, фосфоролиза N-гликозидной связи
нуклеотидов пуриннуклеозидфосфорилазой, последующего дезаминирования
азотисных оснований.
От АМФ и аденозина аминогруппа удаляется гидролитически
аденозиндезаминазой с образованием ИМФ или инозина. ИМФ и ГМФ
превращяются в соответствующие нуклеозиды: инозин и гуанозин под
действием 5’-нуклеотидазы. Пуриннуклеозидфосфорилаза катализирует
расщепление N-гликозидной связи в инозине и гуанозине с образованием
рибозо-1-фосфата и азотистых оснований : гуанина и гипоксантина. Гуанин
дезаминируется и превращяется в ксантин, а гипоксантин окисляется в
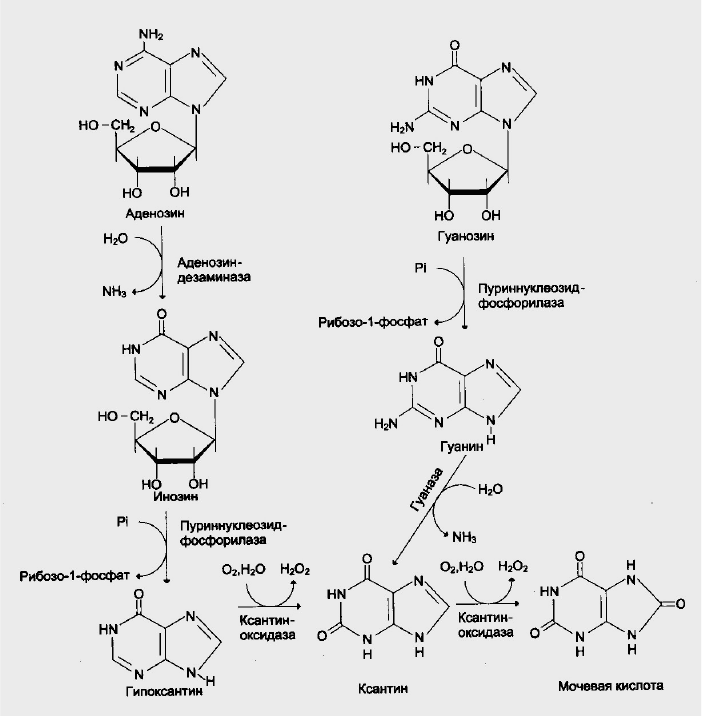
ксантин с помощью ксантиноксидазы, которая катализирует и дальнейшее
окисление ксантина в мочевую кислоту.
Ксантиноксидаза – аэробная оксидоредуктаза, простетическая группа
которой включает ион молибдена, железа (Fе
3+
) и ФАД
+
. Подобно другим
оксидазам, она окисляет пурины молекулярным кислородом с образованием
пероксида водорода. В значительных количествах фермент обраруживается
только в печени и кишечнике.
Рис. 5. Катаболизм пуриновых нуклеотидов до мочевой кислоты.
Мочевая кислота удаляется из организма с мочой и немного через
кишечник. У всех млекопитающие кроме приматов и человека имеется
фермент уриказа, расщепляющая мочевую кислоту с образованием
аллантоина, хорошо растворимого в воде.
Амфибии, птицы и рептилии, подобно человеку лишены уриказы и
экскретируют мочевую кислоту и гуанин в качестве конечных продуктов
обмена.
В сыворотке крови у человека в норме содержание мочевой кислоты
составляет 0,15 – 0,47 ммоль/л или 3-7 мг/дл. Ежесуточно из организма
выводится от 0,4 до 0,6 г мочевой кислоты и уратов.
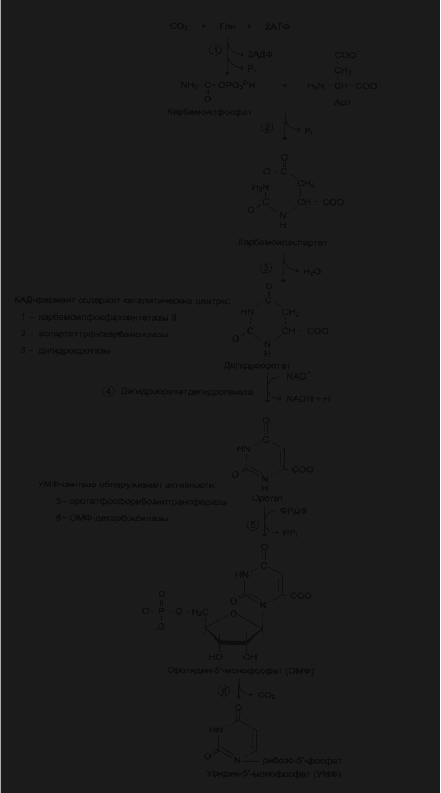
Биосинтез пиримидиновых нуклеотидов
Фонд пиримидиновых нуклеотидов в основном синтезируется de novo
из простых предшественников и только 10-20% от общего количества
образуется по «запасным» путям из азотистых оснований или нуклеотидов.
Пиримидиновое кольцо синтезируется de novo из простых
предшественников: глутамина, СО
2
и аспарагиновой кислоты и затем
связывается с рибозо-5-фосфатом, полученным от ФРПФ.
Процесс протекает в цитозоле клетки. Синтез ключевого
пиримидинового нуклеотида – УМФ идет с участием 3 ферментов, два из
которых полифункциональны.
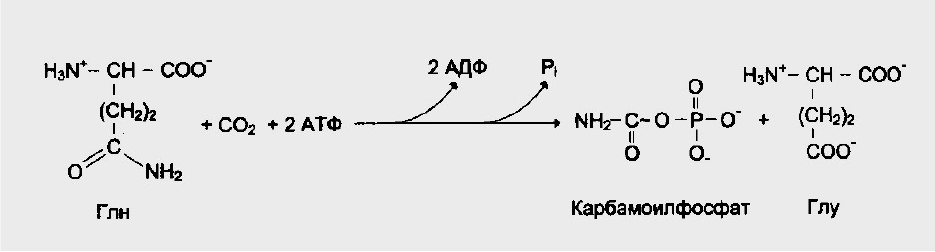
У млекопитающих ключевой, регуляторной реакцией в синтезе
пиримидиновых нуклеотидов является синтез карбамоилфосфата из
глутамина, СО
2
и АТФ, в реакции катализируемой
карбамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле
клетки. В реакции NН
2
-группа карбамоилфосфата образуется за счет амидной
группы глутамина, что отличает эту реакцию от реакции синтеза
карбамоилфосфата в митохондриях в процессе синтеза мочевины из СО
2
,
NH
3
и АТФ с участием КФС I.
Рис.6. Синтез карбамоилфосфата.
Карбамоилфосфат, использующийся на образование пиримидиновых
нуклеотидов, является продуктом полифункционального фермента, который
наряду с активностью КФС II содержит каталитические центры
аспартаттранскарбамоилазы и дигидрооротазы. Этот фермент назвали «КАД-
фермент» - по начальным буквам ферментативных активностей, которыми
обладают отдельные каталитические домены этого белка
Объединение первых трех ферментов метаболического пути в единый
полифункциональный комплекс позволяет использовать почти весь
синтезированный в первой реакции карбамоилфосфат на взаимодействием с
аспартатом и образованием карбамоиласпартата, от которого отщепляется
вода и образуется циклический продукт – дигидрооратат.Отщепляясь от
КАД-фермента, дигидрооратат подвергается дегидрированию NAD-
зависимой дигидроорататдегидрогеназой и превращяется в свободное
пиримидиновое основание – оротовую кислоту или оратат.
Образование УМФ
В цитозоле, оротат становится субстратом бифункционального
фермента – УМФ-синтетазы, которая обнаруживает
оротатфосфорибозилтрансферазную и ОМФ-декарбоксилазную активности.
Первоначально фосфорибозильный остаток от ФРПФ переносится на
оротат и образуется нуклеотид – оротидин-5-монофосфат (ОМФ),
декарбоксилирование которого даёт уридин-5-монофосфат (УМФ)Таким
образом, шесть последовательных реакций синтеза пиримидиновых
нуклеотидов осуществляются тремя ферментами,которые кодируются в
геноме человека тремя различными структурными генами.
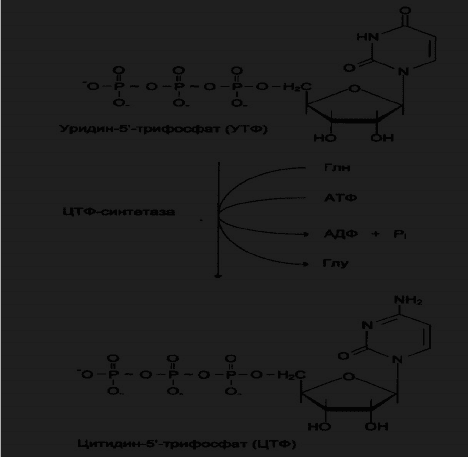
Биосинтез УДФ, УТФ и цитидиловых нуклеотидов
УМФ под действием специфических нуклеозидмонофосфатов (НМФ) и
нуклеизиддифосфатов (НДФ) киназ превращяется в УДФ и УТФ в результате
переноса γ-фосфатного остатка АТФ на соответствующий субстрат.
НМФ-киназа катализирует соответствующую реакцию:
УМФ + АТФ → УДФ + АДФ,
а НДФ-киназа УДФ + АТФ → УТФ + АДФ.
Рис.7. Биосинтез УМФ de novo.
ЦТФ-синтетаза катализирует амидирование УДФ, осуществляя АТФ-
зависимое замещение кетогруппы урацила на амидную группу глутамина с
образованием цитидин-5
/
-трифосфата ( ЦТФ).
«Запасные» пути синтеза пиримидиновых нуклеотидов
Использование примидиновых оснований и нуклеозидов в реакциях
реутилизации препятствует катаболизму этих соединений до конечных
продуктов с расщеплением пиримидинового кольца.
В ресинтезе пиримидинов участвуют некоторые ферменты катаболизма
нуклеотидов.Так, уридинфосфорилаза в обратимой реакции может
рибозилировать урацил с образованием уридина. Превращение нуклеозидов в
нуклеотиды катализирует уридин-цитидинкиназа.
Часть ЦМФ может превращятся в УМФ под действием цитидиндезаминазы и
пополнять запасы уридиловых нуклеотидов.
Рис.8. Синтез ЦТФ из УТФ.
Регуляция синтеза пиримидиновых нуклеотидов
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов
является полифункциональный КАД-фермент. УМФ и пуриновые