Курсовая работа - Малая ядерная РНК
Подождите немного. Документ загружается.

хромосомами образуется достаточное количество белка, кодируемого генами
Х-хромосомы, который активирует промотор главного гена. Транскрипция
главного гена начинается с промотора 1 (на рис. 13 не показан). У самцов с
одной Х-хромосомой количество белка недостаточно, чтобы запустить
главный ген. В результате сплайсинга транскрипта главного гена образуется
РНК 1, содержащая экзоны 1, 4, 5, 6, 7 и 8. В этом случае потенциальные
экзоны 2 и 3 не входят в состав мРНК. Эти события происходят на самых
ранних стадиях развития эмбриона. Немного позднее ген 1 начинает
транскрибироваться как у самок, так и у самцов с использованием другого
промотора 2, лежащего левее. В этом случае 5'-конец РНК-предшественника
будет удлинен, в результате чего экзон 1 сплайсируется иным путем и
становится короче в РНК 1а и 1б по сравнению с РНК 1. Белок 1,
образующийся при трансляции РНК 1, является регуляторным белком
сплайсинга, отсутствующим у самцов. Этот белок направляет сплайсинг у
самок с образованием зрелой РНК 1а, содержащей фрагмент экзона 1, к
которому пришиваются последующие экзоны: 2-4-5-6-7-8. Белок,
определяющий такой характер альтернативного сплайсинга, отсутствует у
самцов. В результате в ядрах клеток развивающегося эмбриона, обладающего
только одной Х-хромосомой, предшественник РНК созревает с образованием
РНК 1б, включающей экзон 3.
Регуляторный белок 1 сплайсинга, образовавшийся у самок на РНК1,
позволяет машине сплайсинга "узнавать" в клетках самок экзон 3 как интрон
и удалять его с образованием РНК1 и РНК1а. Оказывается, экзон 3 содержит
так называемый стоп-кодон, который не кодирует аминокислоты и
останавливает рост полипептидной цепи белковой молекулы. В результате
синтез белка на РНК 1б если и начнется, то быстро оборвется с образованием
очень короткого, распадающегося белкового фрагмента. Таким образом,
"самцовый" тип сплайсинга приводит фактически к инактивации гена,
поскольку ген не способен кодировать образование функционирующей
белковой молекулы. Следовательно, результатом альтернативного
сплайсинга может быть не только образование изоформ белков, кодируемых
одним геном, но и инактивация гена ("главного гена" у самцов). Белок 2,
образующийся у самок, не только постоянно направляет ход сплайсинга с
образованием РНК 1а, но и регулирует созревание РНК 2, содержащей
экзоны 1-2-3. Эта РНК 2 образуется уже на другом гене, входящем в систему
генов определения пола у дрозофилы. У самцов образуется несколько иной
вариант РНК 2 - РНК 2а, включающая несколько увеличенный экзон 2. Этот
фрагмент экзона у самок узнается как интрон. Опять "самцовая" часть экзона
2 содержит стоп-кодон, останавливающий трансляцию, тогда как у самок
образуется активный белок 3, приводящий к образованию РНК 3,
включающей экзоны 1-2-3-4. При отсутствии белка 3 предшественник РНК 3
в ядрах клеток самцов "сплайсируется" таким образом, что
последовательность, соответствующая экзону 4 самок, выглядит как интрон,
а к экзону 3 пришиваются экзоны 5 и 6. На этот раз РНК 3а (экзоны 1-2-3-5-
6) у самцов, кодируемая геном "двойной пол", не содержит вредных стоп-
кодонов, она кодирует функциональный белок. Мишенями действия
21
регуляторных белков, синтезированных на матричных РНК 3 и 3а, являются
гены, дальнейшая работа которых полностью исключает соответственно
развитие самца или самки (рис. 13). Отметим, что, несмотря на вариации в
характере сплайсинга, всегда по краям интронов должны находиться
упомянутые выше канонические нуклеотидные последовательности,
используемые при катализе того или иного пути сплайсинга. Мы проследили
каскад регулируемых событий альтернативного сплайсинга. У самцов он
дважды приводил к образованию дефектной мРНК, не способной кодировать
достаточно длинный и функционирующий белок. У самок наблюдается
последовательное, поэтапное образование матриц РНК для кодирования
белка, регулирующего транскрипцию и/или ход событий альтернативного
сплайсинга.
Примеры альтернативного сплайсинга новообразованной РНК показывают,
как в конечном счете окупаются затраты, в том числе энергетические,
идущие на синтез избытка участков РНК, которые затем удаляются и не
используются для синтеза белка. Альтернативный сплайсинг обеспечивает
регуляцию работы гена, дифференцировку тканей и развитие организма.
Однако биологическая роль прерывистой экзон-интронной структуры генов и
роль интронных последовательностей на этом не заканчивается. Недавно
было обнаружено, что вырезаемые интроны могут быть предшественниками
особых малых ядерных РНК, участвующих в созревании рРНК.
22
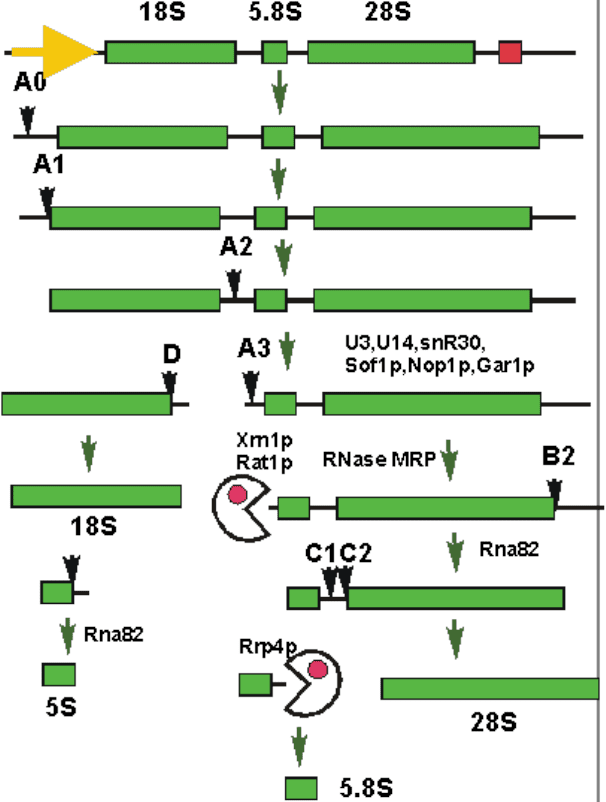
Созревание рРНК. мяоРНК.
Рибосомные РНК синтезируются в виде большого предшественника. Этот
предшественник нарезается на зрелые молекулы рРНК с помощью эндо- и
экзонуклеаз. Кроме того, отдельные нуклеотиды рРНК модифицируются.
Предшественник рРНК прокариот содержит фрагменты, соответствующие
16S, 23S и 5S рРНК. Как правило, в этом транскрипте содержатся также
несколько пре-тРНК. Большие рибосомальные РНК образуют домены
подобного транскрипта. Их 5’ и 3’-концы сближены за счет
комплементарности прилегающих участков РНК. Такая двухцепочечная
структура разрезается РНКазой III, а оставшиеся несколько лишних
нуклеотидов отрезаются экзонуклеазами. Модификация оснований рРНК
проводится набором ферментов, причем один фермент может
модифицировать от 1 до 3 нуклеотидных остатков.
Рис.14. Схема процесса разрезания предшественника рРНК эукариот.
w
У эукариот созревание также включает в себя разрезание предшественника
(рис. 14), причем кроме 18S, 28S, и 5S рРНК появляется еще и 5.8S рРНК.
Кроме различных эндо- и экзонуклеаз, в созревании принимают участие
малые ядрышковые РНК (мяоРНК). Эти РНК локализованы
23

преимущественно в ядрышке – участке ядра, где происходит транскрипция
генов рРНК и сборка пре-рибосом.
Если нарезание предшественника рРНК, в общем, сходно у про- и эукариот,
то система модификации оснований рРНК у эукариот уникальна. В отличии
от прокариот, у высших организмов модифицировано гораздо больше
нуклеотидных остатков рРНК. Среди модификаций подавляющее
большинство это псевдоуридилирование и 2’-O-метилирование рибозы.
Невозможно представить, чтобы модификации всех этих остатков
осуществлялись каждая своим ферментом.
Оказалось, что в ядрышке, помимо мяоРНК, участвующих в разрезании пре-
рРНК, существует множество других мяоРНК. Эти небольшие РНК можно
разделить на 2 класса: C/D и H/ACA (рис. 15).
Рис. 15. Свойства мяоРНК различных классов.
w
МяоРНК обоих классов транскрибируются РНК пролимеразой II, но не как
самостоятельные гены, а в интронах других генов. Часто мяоРНК входят в
состав интронов мРНК рибосомных белков. Есть случаи, когда единственной
функцией некоторых генов служит производство мяоРНК, содержащихся в
интронах. При этом открытая рамка считывания зрелой мРНК прервана
многочисленными стоп-кодонами и не кодирует какого-либо белкового
продукта.
Характерная особенность мяоРНК – короткие участки комплементарности
различным областям рРНК. У РНК C/D класса эта область
24
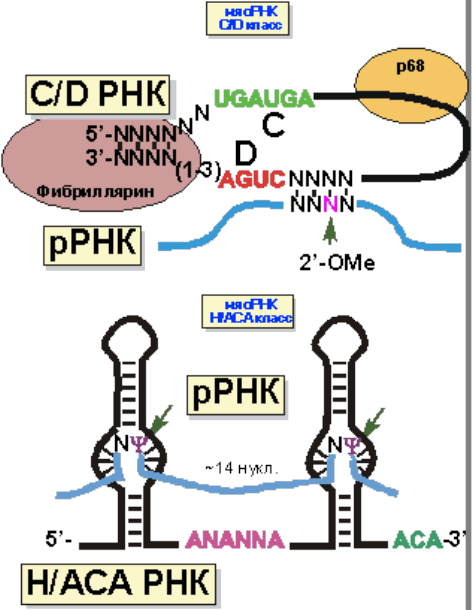
комплементарности более 10 нуклеотидов, а у H/ACA – меньше. С каждым
классом мяоРНК связан свой набор белков. мяоРНП C/D класса
осуществляют 2’-O-метилирование, а H/ACA – псевдоуридилирование рРНК.
Структура мяоРНК представлена на рисунке 17. 5’- и 3’-концы C/D мяоРНК
спарены. Следом расположены две консервативные последовательности
нуклеотидов, из-за которых этот класс и получил свое название. Сразу за D
последовательностью идет участок комплементарности рРНК. Третий
нуклеотид рРНК от участка начала комплементарности 2’-O-метилируется. У
H/ACA мяоРНК также имеется две консервативные последовательности,
давшие название этому классу. Нуклеотиды, комплементарные рРНК
располагаются в двух внутренних шпильках. Остаток уридина в положении,
отмеченном на рисунке 16, изомеризуется в псевдоуридин.
Рис. 16. Модель структуры комплексов
мяоРНК-рРНК, в которых происходит
модификация нуклеотидов.
w
С помощью мяоРНК эукариотические организмы решают проблему
специфичной модификации остатков рРНК, без привлечения огромного
числа специфичных ферментов. Кстати, малые РНК, подобные мяоРНК
участвуют в модификации малых ядерных РНК.
25
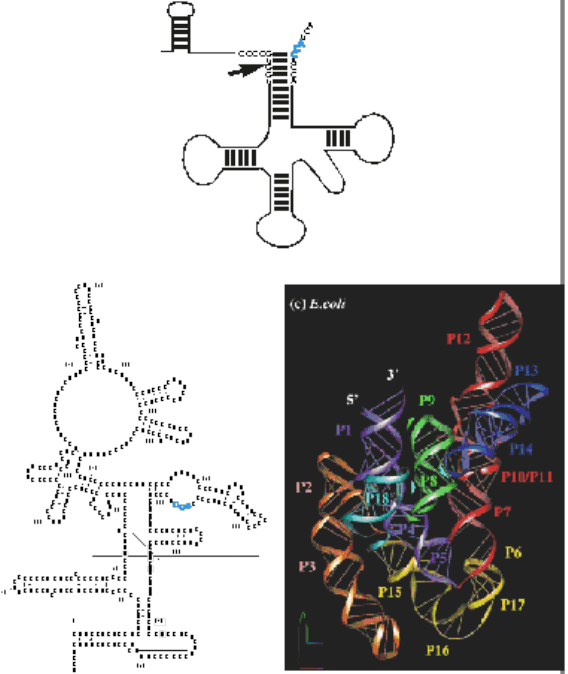
Созревание тРНК
Предшественник тРНК содержит дополнительный фрагмент на 5’-конце
(рис. 17). Этот фрагмент отрезается РНКазой P – рибонуклеопротеином,
основную каталитическую роль в котором играет РНК. РНК-компонент
РНКазы P это рибозим. Белок (или, в случае эукариот, белки) в РНКазе P
играет лишь структурную роль. Также, как и для других рибозимов, РНК
РНКазы P имеет сильную вторичную структуру. В бактериальной РНКазе P
есть участок, комплементарный CCA концу тРНК. Эукариотическая РНКаза
P узнает другие элементы предшественника тРНК.
Рис. 17. Модель вторичной и третичной структур
бактериальной РНКазы P и ее субстрат –
предшественник тРНК.
Формированием 3’-конца молекулы тРНК занимаются несколько белков –
РНКаз. Кроме того, особый фермент – тРНК нуклеотидилтрансфераза
достраивает CCA хвост тРНК, если он был утрачен. У эукариот CCA
последовательность вообще не кодируется в генах тРНК, а добавляется пост-
транскрипционно.
26
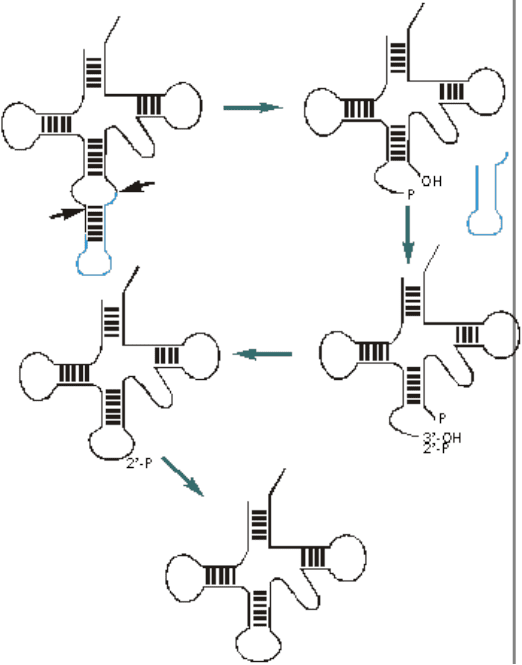
В тРНК эукариот, например дрожжей, есть интроны. В отличии от интронов,
встречающихся в мРНК, они не ограничены какими-либо консервативными
последовательностями и встроены в строго определенный участок тРНК –
антикодоновую петлю (рис. 18).
Рис. 18. Сплайсинг пре-тРНК дрожжей.
w
Предшественник тРНК имеет в районе антикодоновой шпильки
симметричную структуру, узнающуюся димером специфической
эндонуклеазы. Эта нуклеаза вырезает интрон, оставляя фосфат на 3’-конце
5’-концевого фрагмента. Для успешного лигирования фрагментов тРНК,
необходим 3’-гидроксил и 5’-фосфат. Чтобы достичь этого, ферменты,
осуществляющие сплайсинг тРНК, сначала переносят фосфат с 3’-положения
на 2’. Затем проводится фосфорилирование 5’-гидроксила. После
лигирования образуется ковалентно замкнутая тРНК, содержащая
дополнительный фосфат, присоединенный к 2’-OH. Он удаляется
фосфатазой.
27
Заключение.
В заключении хотелось бы выделить основные положения, которые показал
анализ литературных данных.
1. Главной функцией мяРНК является участие в процессе "созревания"
клеточных РНК.
2. Процесс "созревания" клеточных РНК сопровождается химическими
модификациями РНК-предшественника и укорочением его размеров.
Созревание представляет собой регулируемый многостадийный путь, по ходу
которого осуществляется взаимодействие новообразованного предшественника
РНК с белками и другими РНК, обеспечивающими в конце концов образование
функционально активной молекулы РНК. Регуляция процесса созревания РНК -
это способ регуляции активности гена, кодирующего данную молекулу РНК.
3. При созревании информационной РНК имеет место каталитический
процесс удаления из РНК интронов и объединения экзонов, кодирующих
отдельные участки белка. Совокупность этих химических реакций носит
название сплайсинга. Сплайсинг проходит в специальной внутриядерной
многокомпонентной структуре - сплайсосоме, включающей десятки белков и
набор так называемых малых ядерных РНК, обеспечивающих сплайсинг.
4. Молекулярный механизм сплайсинга осуществляется благодаря
комплементарным взаимодействиям нуклеотидов РНК-предшественника и
малых ядерных РНК. Эти взаимодействия играют первостепенную роль в
катализе сплайсинга. Образующиеся сложные структуры определяют точность
сплайсинга. Кроме того, велика роль белков, способных специфично "узнавать"
не только участки РНК, но и друг друга. Сплайсинг определяется
последовательными упорядоченными взаимодействиями компонентов машины
сплайсинга с предшественниками РНК. Исследование молекулярных
механизмов сплайсинга показывает, что каталитическая активность,
определяющая удаление интронов и объединение экзонов, может иметь место и
при отсутствии белков, то есть определяться взаимодействиями участков РНК,
приводящих как к разрыву, так и к воссоединению межнуклеотидных связей.
Нарушение точности сплайсинга при образовании информационных РНК,
вызванное мутацией, может препятствовать трансляции и образованию белка.
5. Многокомпонентность машины сплайсинга позволяет регулировать
образование мРНК за счет изменения концентраций отдельных компонентов
или химической модификации, меняющей биологическую активность молекул,
определяющих сплайсинг.
6. Изменение характера сплайсинга экзонов в разных тканях одного и
того же РНК-предшественника может приводить к образованию разных РНК,
содержащих разные наборы экзонов (альтернативный сплайсинг). В результате
РНК, транскрибируемые с одного гена, будут кодировать белки с разными
свойствами. Выбор путей сплайсинга РНК-предшественника - это способ
регуляции активности генов в разных клетках и тканях организма.
28
Список используемой литературы.
1. Гвоздев В.А. Механизмы регуляции активности генов в процессе
транскрипции // Соросовский Образовательный Журнал. 1996. № 2. С.
22-31.
2. Велц Р., Шмидт К., Мюллер С. Моделирование и синтез рибозимов //
Молекулярная биология - 2000 - т.34 - вып.6 – с.1090-1095
3. Зверева М.Э., Шпанченко О.В. Структура и функции тмРНК (10Sa
РНК) // Молекулярная биология – 2000 – т.34 – вып.6 – с. 1081-1088
4. Кленов М.С., Гвоздев В.А. Формирование гетерохроматина:
ролькоротких РНК и метилирования ДНК // Биохимия – 2005 – т.70 –
вып.11 – с.1445
5. Тайны маленьких РНК // Вокруг света – 2007 - №3
6. Лекции, представленые на сайте кафедры химии природных
соединений МГУ.wwСергиев П.В. Процессинг. 2002 г
http://nature.chem.msu.su/study_lectures_editing_lectureonline_ru.html
7. Томас Р. Чек РНК – фермент // В мире науки. 1987 №1
29