Хант Р.В.Г. Цветовоспроизведение
Подождите немного. Документ загружается.


но не световой поток на единицу телесного угла). Существует множество доказа
тельств тому, что взаимоотношения эти нелинейны.
35.5.1 Корнеквадратная функция
Если колбочковый отклик принять как пропорциональный корню квадратному от
интенсивности стимула, то кривизну линий постоянного цветового тона на диаграмме
цветностей можно предсказать довольно легко (Hunt, 1982). Правда, применение кор
неквадратной зависимости влечет за собой редукцию динамического диапазона сигна
лов, передаваемых из сетчатки в мозг, к примеру: падение интенсивности стимула в
1000 раз ведет к падению величины колбочкового отклика всего в 32 раза.
Однако простые корнеквадратные взаимоотношения касаются далеко не всех воз
можных интенсивностей стимула: когда интенсивность очень мала, т.н. физиологиче
ский шум зрительной системы препятствует извлечению полезной информации из
сверхмалого колбочкового отклика; когда интенсивность стимула очень высока, то от
клик доходит до максимального уровня (насыщение колбочек), за которым уже не в
состоянии меняться (Baylor, 1987). Данные ограничения иллюстрируют неспособ
ность нашей зрительной системы распознавать хроматические модуляции в очень
темных объектах, а также тенденцию к обесцвечиванию особо ярких стимулов (к при
меру, когда раскаленная спираль лампы накаливания рассматривается через окра
шенный фильтр, она все равно выглядит белой либо почти белой).
35.5.2 Гиперболическая функция
Несмотря на сказанное в предыдущем подразделе, командой разработчиков
CIECAM97s вместо корнеквадратной была выбрана гиперболическая функция кол
бочкового отклика (верность такого выбора доказывают физиологические исследова
ния Boynton и Whitten, 1970, а также Valeton и Van Norren, 1983). Отклики колбочек
трех разных типов при данном статусе адаптации зрения сформулированы как:
[]
[]
f
f
n
n
() ( )
() ( )
..
..
rrr
ggg
+= + +
+= +
140 2 1
140 2
073 073
073 073
[]
+
+= + +
1
140 2 1
073 073
f
n
() ( )
..
rbb
,
где «+1» представляет физиологический шум.
Диапазон откликов в этом случае простирается от 1 (минимальное значение) до 41
(максимальное).
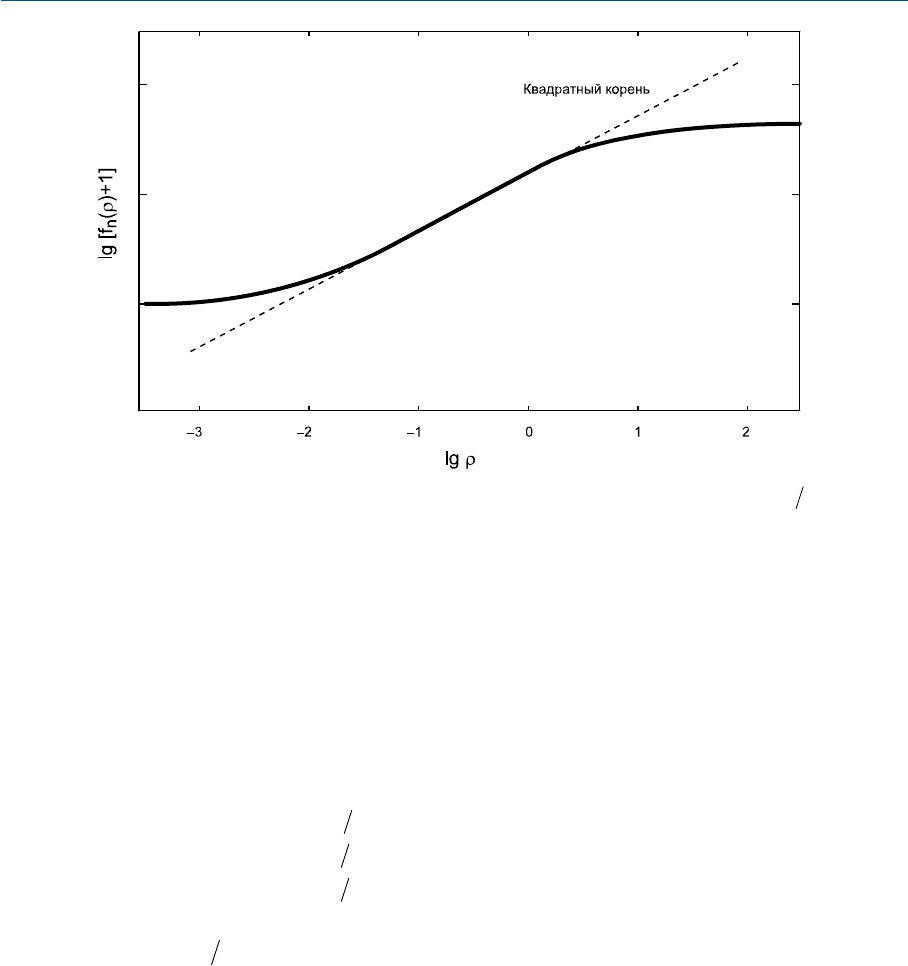
На рис. 35.2 десятичный логарифм
[() ]f
n
r+1
дан как функция от
lgr
. Мы видим, что
в районе центральной части графика отклик длинноволновочувствительных колбочек
устойчиво аппроксимирует корнеквадратную зависимость (пунктирная линия), но на
периферии функция сильно отклоняется от корнеквадратной. Отклики
f
n
()g+1
и
f
n
()b+1
представлены аналогичными кривыми.
35.6 ЯРКОСТНАЯ АДАПТАЦИЯ
Фактический колбочковый отклик зависит не только от интенсивности стимула,
но также от статуса адаптации зрительной системы наблюдателя. Адаптация дает час
тичную (а иногда и полную) компенсацию визуальных эффектов, возникающих в пер
791
ГЛАВА 35 МОДЕЛЬ CIECAM97S
вый момент после смены интенсивности и цветности освещения, являясь физиологи
ческим субстратом такого феномена, как цветовая константность (см. гл. 34).
Мы помним, что учет факта компенсации зрением изменения в цветности (но не ин
тенсивности) освещения выполняется с помощью CAT97 (расчет смены хроматиче
ской адаптации — см. раздел 34.5). Для учета же компенсации смены интенсивности
освещения в CIECAM97s введен специальный коэффициент — коэффициент степени
яркостной адаптации — F
L
. Таким образом, т.н. постадаптационные колбочковые от
клики формулируются так:
rrr
ggg
bbb
anL wr
anL wr
anL wr
=+
=+
=+
fF
fF
fF
()
()
()
1
1
1
,
где
[]
fI I I
n
() ( )
..
+= +140 2
073 073
, а коэффициенты r
wr
, g
wr
и b
wr
— это r, gиb значения
колбочковых откликов на согласованный стимул в эталонных условиях просмотра
и при эталонном равноэнергетическом белом (S
E
). Эталонному белому стимулу, кото
рый создает идеальный отражающий рассеиватель, присвоена эталонная цветность,
равная цветности равноэнергетического S
E
осветителя (эталонная цветность). Однако
заметим, что Yзначение (100), присвоенное эталонному белому стимулу (Y
wr
), не ока
зывает влияния на XYZзначения стимула согласованного. Так происходит потому,
что параметры эталонного белого стимула (R
wr
, G
wr
и B
wr
), участвующие в расчете, полу
чены из отношений между его трехстимульными значениями, что равно отношениям
между координатами его цветности.
Однако когда прогнозируется числовой коррелят светлоты (L) исследуемого стиму
ла, Yзначение эталонного белого влияет на результат (см. определение светлоты в раз
деле 7.2), и белый, принятый в эталонных условиях, подбирают как согласованный
тестовому белому. Идеальный отражающий рассеиватель не очень подходит для реше
792
ГЛАВА 35 МОДЕЛЬ CIECAM97S
Рис. 35.2 Логарифмированный колбочковый отклик (
lg[ ( ) ]f
n
r+1
, где
[]
f
n
() ( )
..
rrr=+40 2
073 073
)
как функция от логарифма эффективно поглощенного излучения (
lgr
).
ния такой задачи, поскольку если с его помощью оценивать ощущение светлоты от
тестового белого стимула, то светлота последнего окажется ощутимо меньшей — то
есть ощущение окажется сероватым, но не подлинно белым. Отметим также, что вы
бор Yзначений эталонного и тестового белых влияет на показатели их субъективной
яркости (Qвеличины) при эталонном статусе адаптации.
Когда коэффициенты r
wr
, g
wr
и b
wr
используются без применения F
L
, это значит, что
компенсация зрением изменений в уровне освещенности объявляется полной. Однако
на практике компенсация эта, как правило, неполна: вспомним, что при слабом осве
щении объекты воспринимаются менее яркими (с меньшей субъективной яркостью),
чем при освещении сильном, даже если сетчатка при этом полностью адаптирована.
1
Итак, коэффициент F
L
моделирует неполноту яркостной адаптации и определен
уравнением:
FkL kL
LA A
=+-02 5 011 5
442
3
.( ).( )( )
,
где L
A
— фотометрическая яркость адаптирующего поля, а
kL=+15 1()
A
.
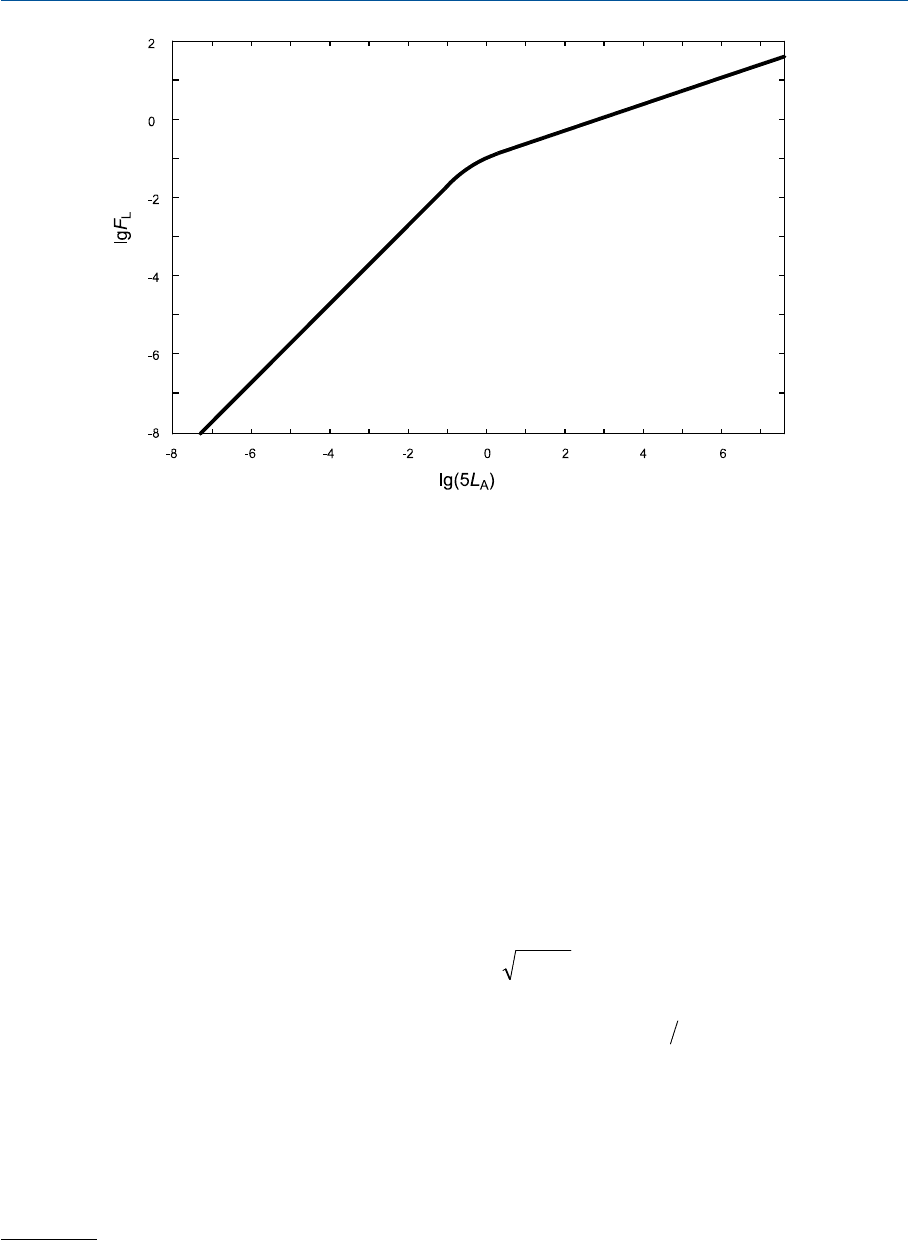
На рис. 35.3 десятичный логарифм
F
L
дан как функция от
lg( )5L
A
: четко видно, что
при фотопических уровнях освещенности сцены (
515 0LL
AA
>>;lg( )
) F
L
примерно про
порционален корню кубическому из
5L
A
(тангенс угла наклона кривой равен 1/3), что
имитирует неполноту яркостной адаптации при изменении фотометрической яркости
адаптирующего поля. Полную компенсацию можно имитировать, если
F
L
const=
.
На скотопических уровнях освещенности (
5015 1LL
AA
<<-.; lg( )
) F
L
примерно про
793
ГЛАВА 35 МОДЕЛЬ CIECAM97S
Рис. 35.3 Десятичный логарифм коэффициента степени яркостной адаптации как функция от
десятичного логарифма пятикратной фотометричиеской яркости адаптирующего поля
(т.е. фотометрической яркости типичного белого стимула).
1
Но при этом могут оставаться равновелико светлыми по отношению к какомулибо стиму
лу, субъективная яркость которого принята за эталонную. — Прим. пер.

порционален
5L
A
(тангенс наклона кривой 1), что имитирует полное отсутствие ярко
стной адаптации.
Наконец, отметим, что фотометрическая яркость белого стимула при данном ста
тусе адаптации равна пятикратной фотометрической яркости адаптирующего поля.
35.7 КРИТЕРИИ АХРОМАТИЧНОСТИ ПРИ ПОСТОЯННОМ
ЦВЕТОВОМ ТОНЕ
Как было показано в разделе 2.3, тройственный отклик колбочек обрабатывается
нейронами сетчатки. Результатом обработки является цветоразностный (оппонент
ный) сигнал, отправляемый по волокнам зрительного нерва в высшие отделы головно
го мозга. Оппонентные сигналы можно представить как:
C
C
C
1
2
3
=-
=-
=-
rg
gb
br
AA
AA
AA
Однако, разумеется, существуют и три дополнительных варианта:
¢
=-
¢
=
¢
=-
C
C
C
1
2
3
gr
b-g
rb
AA
AA
AA
,
но для простоты мы ограничимся лишь первыми тремя.
Напомним, что те стимулы, в восприятии которых отсутствует цветовой тон (бе
лые, серые и черные), называются ахроматическими. Как было показано в разделе
2.5, критерием ахроматичности стимула является равенство колбочковых откликов
на него:
rgb
aaa
==
,
и, следовательно, —
CCC
123
0===
Понятно, что чем более величины С
1
, С
2
и С
3
удаляются от нуля, тем выше полнота
цвета стимула.
Понятно также, что критерием постоянства цветового тона является:
CCC
12 3
=const
35.8 ЭФФЕКТЫ ЯРКОСТНОЙ АДАПТАЦИИ
На рис. 35.4 величины r
a
даны как функция от
lg( ) lg5LI
Aw
rr =
, где r
w
— это от
клик rколбочек на тестовый белый стимул. Если
5L
A
— это фотометрическая яркость
белого стимула, а тестируемый образец при этом имеет ту же цветность, то выражение
5L
Aw
rr
будет равно фотометрической яркости образца. Кривые рис. 35.4 даны по
794
ГЛАВА 35 МОДЕЛЬ CIECAM97S
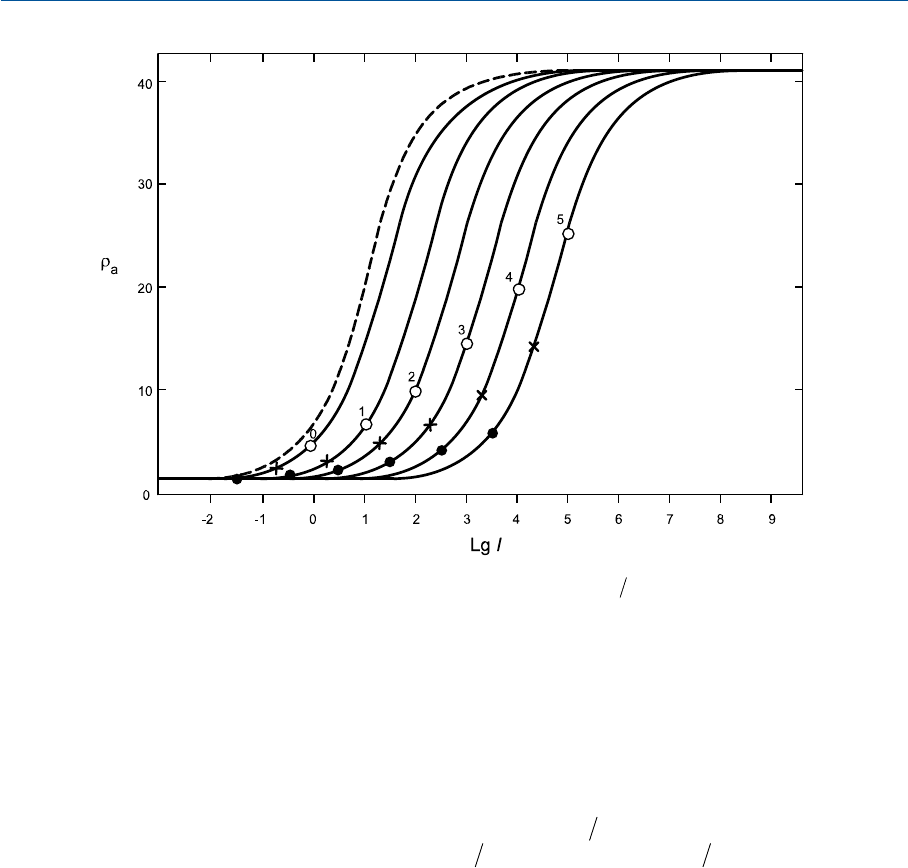
значениям
lg( )5L
A
, равным 5, 4, 3, 2,1и0(сплошные линии), а также при темновой
адаптации (пунктирная линия).
Давайте рассмотрим кривую «3», относящуюся к
lg( )5L
A
= 3: белый кружок на кри
вой относится к стимулу, имеющему то же значение
rr
w
, что и у тестового белого,
а черный кружок — к стимулу, по которому
rr
w
равно 0.03162 от
rr
w
по тестовому бе
лому (что на 1.5 единицы ниже 3 по логарифмической шкале). Позиция адаптирующе
го стимула (стимула, создаваемого адаптирующим полем) — L
A
, фотометрическая яр
кость которого равна 1/5 (20% — на 0.7 ниже 3 по логарифмической шкале) фотомет
рической яркости белого, обозначена крестиком. При этом сразу отметим, что анало
гичные рассуждения применимы к g
a
и b
a
.
Следовательно, когда
lg( )5L
A
= 3, тестовый белый оказывается представлен про
зрачным кружком в одной и той же позиции на всех трех rgbкривых, а стимул с той
же цветностью и с фотометрической яркостью, составляющей 0.03162 от яркости тес
тового белого, — черным кружком в одной и той же позиции на всех трех графиках.
Таким образом, участок кривой «3» между прозрачным и черным кружками пред
ставляет диапазон стимулов между белым (точнее, белым при данном статусе адапта
ции — т. е. тестовым белым) и черным, фотометрическая яркость которого составляет
3.162%от фотометрической яркости тестового белого (когда
lg( )5L
A
= 3 логарифмиро
ванным кд/м
2
).
Остальные кривые на рис. 35.4 представляют аналогичные диапазоны стимулов
при прочих величинах
lg( )5L
A
: мы видим, что диапазоны эти прогрессивно уменьша
795
ГЛАВА 35 МОДЕЛЬ CIECAM97S
Рис. 35.4 Отклики rколбочек как функции от LgI (где
IL=5
A
w
rr
,аL
A
— фотометрическая
яркость адаптирующего поля в кд/м
2
) при пяти уровнях
lg( )5L
A
, равных 5, 4, 3, 2, 1 и 0 лога
рифмированных кд/м
2
(сплошные линии), а также при темновой адаптации (пунктирная ли
ния). Белые кружки: тестовый белый. Черные кружки: 3.162% черного. Крестики: адаптирую
щее поле (фотометрическая яркость равна 1/5 от яркости тестового белого).

ются в левом направлении и растут в правом. Отметим, что физиологи в своих отчетах
приводят целые семейства аналогичных кривых (Valeton и Van Norren, 1983).
Sобразная форма обсуждаемых кривых свидетельствует о том,—авконтексте мо
делирования результатов цветового восприятия — прогнозирует то, что полнота цвета
стимулов одной и той же цветности падает по мере падения коэффициента их фотомет
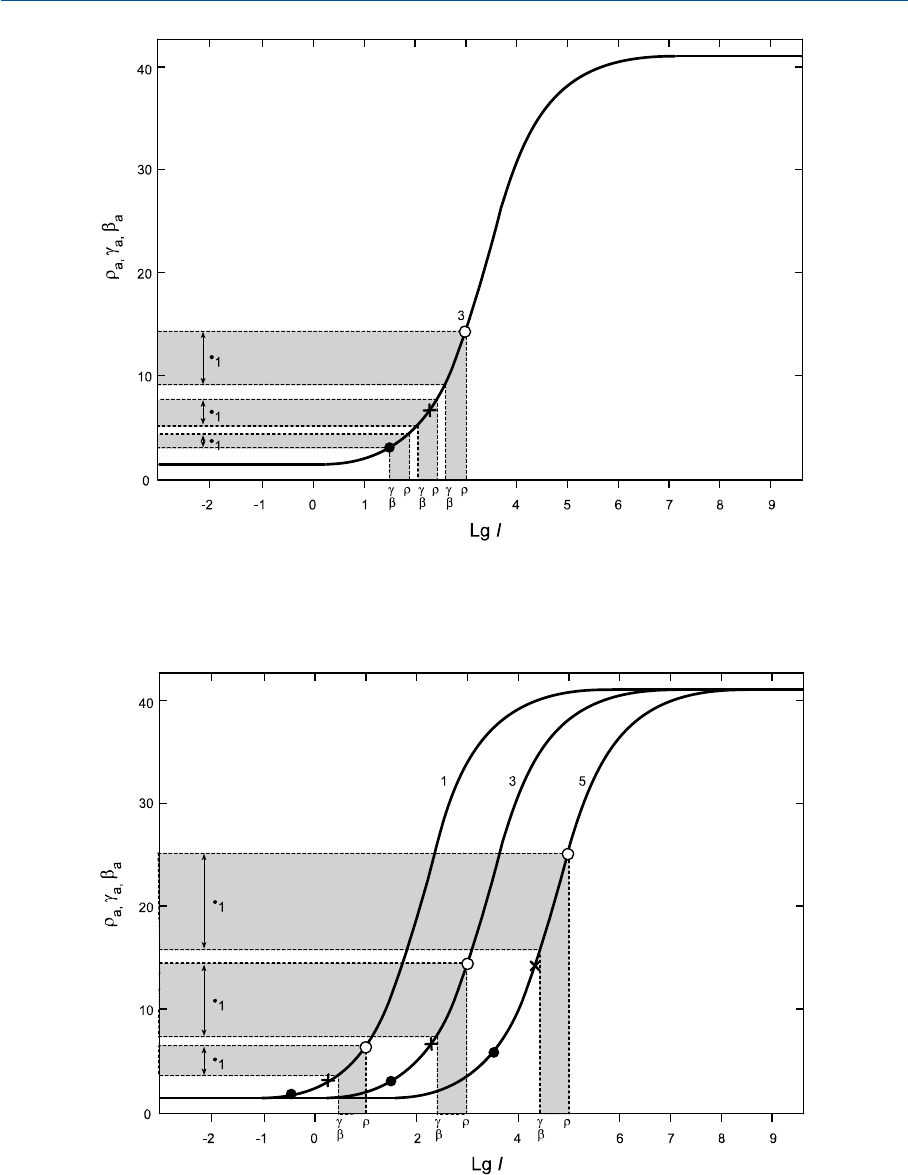
рической яркости (и наоборот). Действительно, если обратиться к рис. 35.5, то мы вновь
увидим кривую «3», но с вертикальными пунктирными линиями, относящимися к
трем красным стимулам одной общей цветности, но разных фотометрических яркостей.
О постоянстве цветности этих стимулов свидетельствуют одинаковые расстояния (0.5)
по оси
lg I
между позициями колбочковых поглощений излучения (r>gи b; g = b).
По мере снижения фотометрической яркости позиции на
lg I
оси смещаются влево.
Исходом является смещение позиций откликов в участок кривой с меньшим накло
ном. Следовательно, отличие между r
a
иg
a
откликами снижается, и, разумеется, сни
жается оппонентный сигнал С
1
(С
3
будет снижаться аналогично), что, как было показа
но ранее, влечет за собой снижение полноты цвета стимула. Таким образом, если при
данной цветности стимула падает его фотометрическая яркость — падает и полнота
цвета этого стимула (что полностью подтверждается практикой).
1
Вернемся к рис. 35.4. Если фотометрическая яркость адаптирующего стимула
2
(L
A
)
падает, кривые смещаются влево, указывая на рост чувствительности сетчатки. Одна
ко последнее не может обеспечить полной компенсации, и, следовательно, падает
субъективная яркость стимула тестируемого. Мы видим также, что позиции точек,
представляющих тестовый белый стимул (¡), адаптирующий (´) и черный (l) плавно
смещаются в область меньшего наклона кривой. Сие ведет к сокращению отличий в от
кликах на белый, адаптирующий и черный стимулы. У хроматичных стимулов при
этом, напомним, падает их полнота цвета.
Сказанное выше проиллюстрирует рис. 35.6, на котором даны кривые 1, 3 и 5 из
рис. 35.4: вертикальные линии в точке пересечения с кривой «5» указывают на то, ка
кова величина
lg I
по rотклику на красный стимул при его фотометрической яркости,
равной фотометрической яркости белого (то же по gиbоткликам, на 0.5 логарифми
ческих единиц меньшим). Соответствующее значение оппонентного отклика С
1
дано
слева. Хорошо видно, что у кривых «1» и «3» величины С
1
ощутимо меньше (что также
касается и С
3
оппонентных откликов).
Итак, CIECAM97s прогнозирует эффект потери полноты цвета стимулов по мере
снижения уровня освещенности (что полностью подтверждается практикой).
Из рис. 35.6 также видно, что наклон кривых возле черной точки (l) слабеет по мере
смещения кривых влево. Сие указывает на то, что CIECAM97s эффективно прогнози
рует снижение различимости темных стимулов при тусклом освещении (и что также
полностью подтверждается практикой).
Наконец, отметим, что кривые рис. 35.4, 35.5 и 35.6 демонстрируют также и то,
что по стимулам, фотометрическая яркость которых много больше, чем у тестового бе
лого, колбочковые отклики приближаются к максимально возможному уровню, что
также влечет за собой снижение полноты цвета этих стимулов.
796
ГЛАВА 35 МОДЕЛЬ CIECAM97S
1
Автор скромно умалчивает, что данный феномен именуется «эффектом Ханта» (Fairchild,
2004). — Прим. пер.
2
Адаптирующий стимул — интегральный стимул, созданный адаптирующим полем.
797
ГЛАВА 35 МОДЕЛЬ CIECAM97S
Рис. 35.5 Кривая «3» (из рис. 35.4), на которой даны позиции стимулов одной цветности, но
трех разных фотометрических яркостей. Хорошо видно, что по мере снижения фотометричес
кой яркости стимула оппонетный отклик на него падает.
Рис. 35.6 Кривые «1», «3» и «5» (из рис. 35.4), на которых отмечены позиции стимулов одной
цветности и одной фотометрической яркости при трех разных уровнях яркости адаптирующе
го стимула (допущено, что фотометрическая яркость тестового белого во всех трех случаях
в 5 раз выше яркости адаптирующего поля). Хорошо видно, что по мере снижения яркости
адаптирующего поля оппонентный отклик на исследумый стимул также падает.

35.9 КРИТЕРИИ ОДНОЗНАЧНОСТИ ЦВЕТОВЫХ ТОНОВ
Мы помним, что существуют четыре однозначных цветовых тона: красный, зеле
ный, желтый и синий. CIECAM97s позиционирует эти тона исходя из постоянства от
ношения C
1
к С
2
либо к C
3
(поскольку C
1
+ C
2
+ C
3
= 0, то, ежели одно из этих соотноше
ний постоянно, и остальные тоже будут постоянными, и нет необходимости в дополни
тельной спецификации).
Однозначный красный
С
1
= С
2
Однозначный зеленый
С
1
= С
3
Однозначный желтый
С
1
= С
2
/11
Однозначный синий
C
1
=C
2
/4
На рис. 35.7 сплошными линиями отмечены предикторы, полученные исходя из
данных критериев, по стимулам, предъявляемым на высоких уровнях яркости адап
тирующего стимула (L
A
= 200 кд/м
2
при Cосветителе); в подтверждение эффективно
сти модели приведены пунктирные линии, полученные экспериментальным путем с
использованием системы NCS (см. раздел 8.12).
35.10 КРАСНОВАТОСТЬLЗЕЛЕНОВАТОСТЬ
И ЖЕЛТОВАТОСТЬLСИНЕВАТОСТЬ
Когда речь идет о красных оттенках, условием однозначности красного тона явля
ется C
1
= C
2
. Отклонение от этого условия ведет к появлению в красном либо желтого
чувственного компонента (нарастание желтоватости), либо синего (нарастание сине
ватости). И первое, и второе указывает на усиление неравенства между С
1
и С
2
, то есть
на то, что разность между оппонентными откликами С
1
и С
2
начинает удаляться от
ноля (отметим, что С
1
вычитается из С
2
, чтобы на нарастание желтизны указывали по
ложительные значения).
Аналогично, так как условием однозначности зеленого тона является C
1
= C
3
,нана
растание желтоватости или синеватости в зеленых укажет отклонение C
1
– C
3
от ноля
(при нарастании желтизны в область положительных значений). Следовательно, мера
желтизны или синевы красных и зеленых тонов будет представлять собой среднее
арифметическое между двумя разностями:
CCCC
2113
2
-+-
,
что равно
05
23
.( )CC-
.
Аналогичные рассуждения применимы к красноватости/зеленоватости желтых
и синих стимулов. Однако коль скоро однозначный желтый чувственно оценивается
более интенсивным, чем однозначный синий (Kuehni, 2004), то среднее арифметиче
ское в этом случае неприменимо, и красноватость/зеленоватость желтых и синих сти
мулов выражается как:
CC a
12
11-=(/)
798
ГЛАВА 35 МОДЕЛЬ CIECAM97S
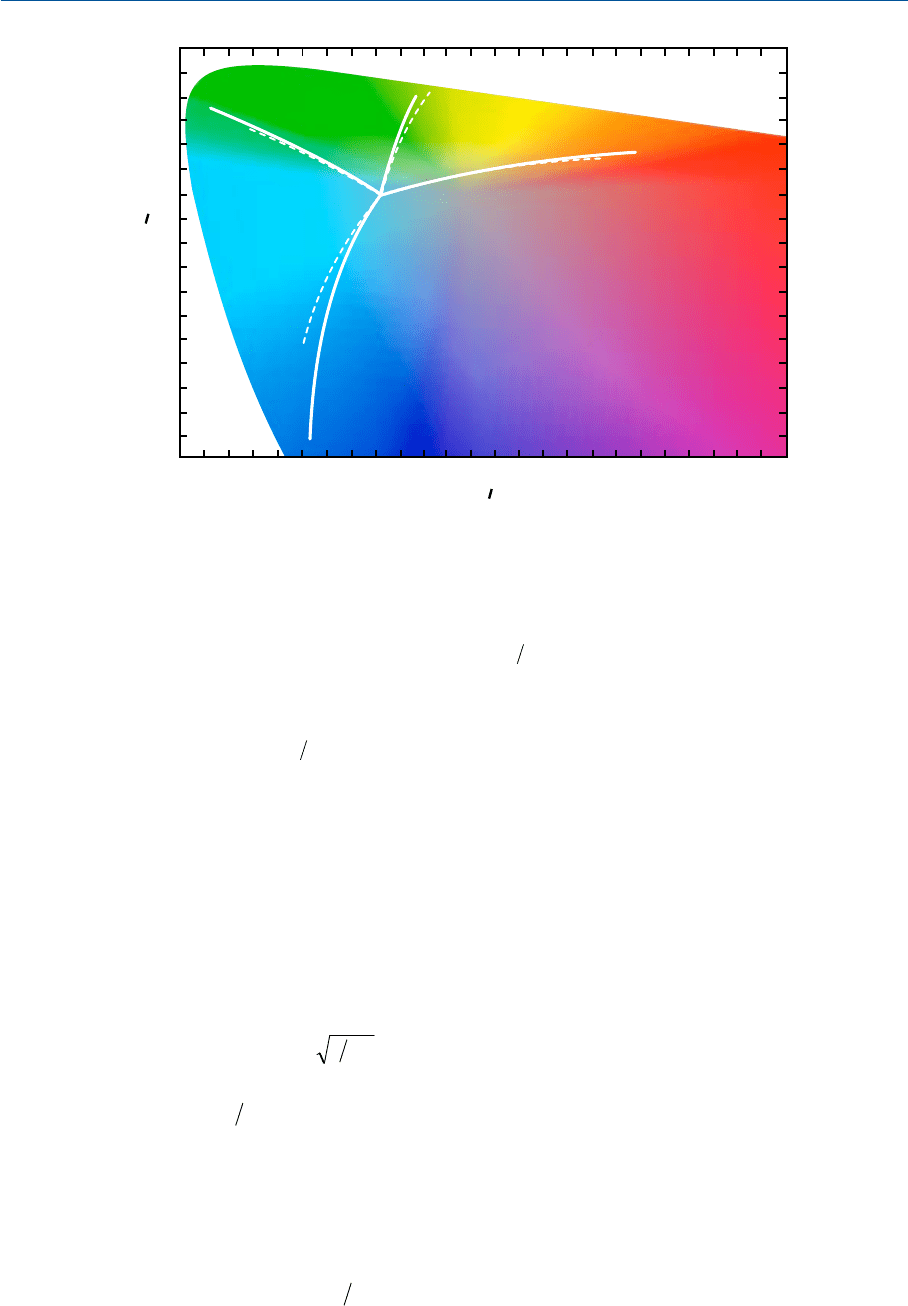
Теперь нам необходимо объединить корреляты желтоватости/синеватости и крас
новатости/зеленоватости, дабы получить метрику цветового тона. Однако прежде на
помним, что общее количество bколбочек в сетчатке составляет всего лишь 1/20 от
суммы rиgколбочек (Warlaven и Bouman, 1966). Если рассуждать в контексте соот
ношения «полезный сигнал/шум», то становится понятным, что желтосиний оппо
нентный сигнал сетчатки должен обладать меньшим весом, нежели сигнал
краснозеленый. С целью учета сего обстоятельства CIECAM97s использует коэффи
циент 1/4.5 (примерно равен
120
), в результате чего желтоватостьсиневатость вы
ражается как:
12 45
23
()/.CC b-=
35.11 УГОЛ ЦВЕТОВОГО ТОНА
Количественным показателем цветового тона (hue) является, как мы знаем, его
угол h:
hba= arctan( )
799
ГЛАВА 35 МОДЕЛЬ CIECAM97S
v
0 0.1 0.2 0.3 0.4 0.5
0.4
0.5
0.6
0.3
u
Y
R
B
G
Рис. 35.7 Сплошные линии: локусы однозначных цветовых тонов, спрогнозированные
CIECAM97s при фотометрической яркости адаптирующего поля 200 кд/м
2
. Пунктирные ли
нии: локусы однозначных цветовых тонов в системе NCS при Стандартном Сосветителе. Что
бы гарантировать «попадание» постадаптационных колбочковых откликов (r
a
, g
a
, b
a
) на линей
ный участок кривой рис. 35.2, выражение
rg b
aa a
++()21 20
положено равным 30. То же выра
жение по идеальному отражающему рассеивателю — 43. Поскольку (как показано на рис.
35.2) гиперболическая функция в своем центральном участке аппроксимирует простую сте
пенную функцию, цветности, соответствующие локусам постоянного цветового тона, лишь
слегка зависимы от
rg b
aa a
++()21 20
, что и гарантирует «попадание» r
a
, g
a
и b
a
в центральный
участок кривой.
По NCS фотометрические яркости были максимально допустимыми системой.

Угол цветового тона ляжет между 0° и 90°, если аиbзначения положительны; ме
жду 90° и 180° — если aзначение отрицательно, а b — положительно; между 180° и
270° — если аиbзначения отрицательны; между 270° и 360° — если aзначение поло
жительно, b — отрицательно.
35.12 КОРРЕЛЯТ ЧИСТОТЫ ЦВЕТА
Мы знаем, что полнота цвета — это перцепционный показатель, являющийся мерой
выраженности цветового тона и, следовательно, мерой желтоватости/синеватости и
красноватости/зеленоватости. Однако, прежде чем (с целью расчета величины корреля
та полноты цвета) мы объединим аиbпоказатели, необходимо поговорить о таких пер
цепционных переменных, как чистота цвета (saturation)инасыщенность (chroma).
35.12.1 Коэффициент эксцентричности
Необходимость в коэффициенте е — т.н. коэффициенте эксцентричности (ориги
нальности) продиктована эксцентричностью позиции ахроматической точки внутри
контуров малой постоянной чистоты цвета (Hunt, 1985). Ахроматическая точка про
грессивно стремится к этим контурам, если цветовой тон стимула меняется от желтого
к красному и от зеленого к синему. Сие указывает на усиление ощущения хроматично
сти по мере изменения цветового тона в направлениях желтый
®
красный и зеленый
®
синий. Чтобы учесть сие свойство восприятия, коэффициенту эксцентричности
присвоены следующие значения по однозначным цветовым тонам, углы которых (по
лученные из
CCC
12 3
) равны:
Красный Желтый Зеленый Синий
h
20.14 90.00 164.25 237.53
e
0.8 0.7 1.0 1.2
Значения e на промежуточных цветовых тонах линейно интерполируются по урав
нению:
ee e e hh h h=+ - - -
121 1 21
()()( )
,
где e
1
и h
1
— это соответственно eиhвеличины однозначных цветовых тонов, имею
щих ближайшее меньшее значение h, а e
2
и h
2
— eиhвеличины однозначных цвето
вых тонов, имеющих ближайшее наибольшее значение h.
На рис. 35.8 e дано как функция от h.
35.12.2 Коэффициент шума
В CIECAM97s введен коэффициент 10/13, учитывающий межканальный шум сис
темы (Hunt, 1982).
35.12.3 Коэффициент хроматической индукции окружения
CIECAM97s задействует также т.н. коэффициент хроматической индукции окруже
ния (N
c
), позволяющий учесть тот факт, что тусклое или темное окружения исследуе
мого стимула могут понизить полноту его цвета.
800
ГЛАВА 35 МОДЕЛЬ CIECAM97S