Емцев В.Т., Мишустин Е.Н. Микробиология: учебник для вузов
Подождите немного. Документ загружается.


ты до кетоглюконовых кислот. Перечисленные превращения осу-
ществляются по пентозофосфатному пути представителями рода
Gluconobacter. Особый интерес представляет окисление уксусно-
кислыми бактериями D-сорбита до L-сорбозы. Последняя требуется
в больших количествах для синтеза витамина С.
Интересно отметить, что представитель рода Acetobacter —
A. xylinum при росте на среде с глюкозой или другими источниками
углерода способен формировать внеклеточную слизистую пленку,
состоящую из чистой целлюлозы. Целлюлозные фибриллы пред-
ставляют собой рыхлую массу, окружающую клетки. В культуре этот
организм образует пленку толщиной 1 см и более, состоящую из
целлюлозы и бактериальных клеток.
Окисление углеводов до лимонной и других органических
кислот. Углеводы могут окисляться до лимонной кислоты и других
органических кислот. При рассмотрении процессов дыхания и бро-
жения было отмечено, что некоторые микроорганизмы не полно-
стью окисляют те или иные органические соединения. В этом слу-
чае происходит накопление продуктов неполного окисления — ок-
салата, цитрата, сукцината, фумарата, малата, аконитата, глюконата
и других кислот. Подобные процессы часто вызывают грибы. Так,
виды родов Rhizopus и Мисоr вызывают окисление углеводов глав-
ным образом до малата, в небольших количествах до сукцината, фу-
марата, малата, ацетата и муравьиной кислоты, а также этанола; ро-
дов Aspergillus и Penicillium — до глюконата, оксалата и цитрата.
Микробиологический синтез цитрата, глюконата, а-кетоглу-
тарата, сукцината, фумарата, малата и ряда других органических
кислот обычно осуществляется при интенсивной аэрации. Перечис-
ленные кислоты служат промежуточными продуктами метаболизма
соединений углерода, в том числе цикла трикарбоновых кислот.
Большое практическое значение имеет микробиологическое
получение цитрата из углеводов с использованием гриба Aspergillus
niger, превращающего почти 60% глюкозы в лимонную кислоту.
Возможность такого процесса была установлена С. П. Костычевым
и В. С. Буткевичем. Сейчас разработан и промышленный способ
микробиологического получения лимонной кислоты, необходимой
в медицине, фармацевтической, пищевой и химической промыш-
ленности, а также при дублении кож, в печатном деле и т. д.
В процессе окисления глюкозы наряду с цитратом всегда об-
разуется глюконат, причем выход последнего зависит от штамма
гриба и от рН среды. Попутно выделяются оксалат и сукцинат.
В России, США и Японии налажено производство итаконо-
вой кислоты на основе деятельности Aspergillus terreus. Итаконовая
кислота — продукт превращения углеводов при участии и другого
гриба рода Aspergillus — A. itaconicus. Итаконовую кислоту применя-
184
ют в производстве синтетических смол, синтетических волокон, ин-
сектицидов, красителей и других веществ.
В последнее время разработан способ получения цитрата из
углеводородов (н-парафинов) с помощью дрожжей рода Candida.
Указанные дрожжи используют также для получения изоцитрата,
а-кетоглутарата, фумарата, малата и сукцината.
9.6. Разложение целлюлозы и других органических
веществ микроорганизмами
В состав целлюлозы входит более 50% всего органического углерода
биосферы, это наиболее распространенный полисахарид раститель-
ного мира, высшие растения на 40—70% состоят из целлюлозы.
В связи с большим количеством синтезируемой в природе целлюло-
зы микроорганизмы, ее разлагающие, играют очень важную роль
в процессе минерализации органического вещества и круговороте
углерода.
Разнообразие микрофлоры, способной разлагать целлюлозу в
почве, позволяет трансформировать это вещество в различных усло-
виях аэрации, в кислой или щелочной среде при низких или высо-
ких влажности и температуре. Для большинства микроорганизмов,
разлагающих целлюлозу, характерна высокая специфичность по от-
ношению к этому веществу. Разложение целлюлозы осуществляют
аэробные микроорганизмы (бактерии и грибы) и анаэробные мезо-
фильные и термофильные бактерии.
Аэробное разложение целлюлозы. Группа аэробных целлю-
лозоразлагающих микроорганизмов наиболее богато представлена в
почве. В 1918 г. X. Б. Хутчинсон и Дж. Клейтон выделили из почвы
целлюлозоразлагающую бактерию, имеющую вид длинной верете-
нообразной палочки с острыми концами. Микроорганизму было да-
но название Spirochaeta cytophaga. Сейчас эту бактерию относят к се-
мейству Cytophagaceae, роду Cytophaga. Впоследствии были описаны
другие виды цитофаг. В настоящее время среди видов этого рода из-
вестны формы, разлагающие хитин (рис. 39).
Бактерии рода Cytophaga очень требовательны к условиям сре-
ды, обычно они в большом количестве встречаются в навозе и поч-
вах, удобренных навозом. То же свойственно и представителям рода
Sporocytophaga, разлагающим целлюлозу. Последние отличаются от
видов рода Cytophaga способностью образовывать микроцисты.
В расщеплении целлюлозы принимают участие миксобактерии
порядка Myxobacteriales, семейств Мухососсасеае (род Myxococcus),
Archangiaceae (род Archangium), Sorangiaceae (роды Sorangium и Poly-
angium), широко распространенные в почвах разных климатических
зон.
185

Рис. 39. Разложение фильтровальной бумаги в местах роста бактерий рода Cytophaga —
слева; целлюлозное волокно, покрытое бактериями, — справа (по: С. Н. Виноградский)
В почвах встречаются представители рода Cellulomonas. Это
аэробные грамположительные подвижные палочковидные бактерии
неправильной формы; с возрастом они иногда превращаются в кок-
ки. Бактерии данного рода участвуют в разложении целлюлозы в
аэробных условиях, однако способны и к анаэробному развитию.
Встречаются эти бактерии в почвах, богатых минеральными форма-
ми азота.
Могут усваивать целлюлозу единичные виды Pseudomonas
(P. fluorescens var. cellulosae) и некоторых других бактерий. Актино-
мицеты и грибы, обитающие в относительно бедных, кислых поч-
вах, в аэробных условиях медленно разрушают целлюлозу. К актино-
мицетам, разлагающим целлюлозу, относятся представители родов
Streptomyces {Streptomyces cellulosae), Streptosporangium, Micromonospora
(Micromonospora chalcea); к грибам — представители родов Fusarium,
Chaetomium, а также отдельные виды — Trichoderma viride, Aspergillus
fumigatus, Botrytis cinerea, Rhizoctonia solani, Myrothecium verrucaria.
В разрушении целлюлозы участвуют и хитридиомицеты, среди кото-
рых много паразитов.
Анаэробное разложение целлюлозы. Большинство предста-
вителей анаэробных целлюлозоразлагающих бактерий, найденных
в природе, относят к семейству Васillасеае, роду Clostridium. Эти ви-
ды обитают в почвах, компосте, навозе, речном иле и сточных во-
дах. Клостридии устойчивы к кислотности и распространены не
только в нейтральных, но и кислых почвах. Типичный представи-
тель рода, участвующий в разложении целлюлозы при температуре
186
30—40 °С, — Clostridium omelianskii. Впервые вид выделен известным
микробиологом В. Л. Омелянским в 1902 г. Этот микроорганизм
имеет палочковидную форму (4—8 х 0,3—0,5 мкм), подвижен. Для
него характерны толстые споры, поэтому спорообразующая клетка
сильно раздувается и становится похожей на барабанную палочку.
Разлагать целлюлозу может и другой мезофильный вид — С. cellobio-
pаrит.
Среди анаэробных целлюлозоразлагающих бактерий, встре-
чающихся в почве, навозе и компостах, есть и термофилы. Они ак-
тивно сбраживают целлюлозу. Так, оптимальная температура для
развития С. thermocellum около 60 °С, биологический максимум при-
ближается к 70 °С; при 40, 45 °С эта бактерия развивается слабо.
К термофильным целлюлозоразлагающим анаэробам относится и
вид Thermoanaerobacter ethanolicus, выделенный из горячих источни-
ков. Мезофильные и термофильные анаэробные бактерии хорошо
используют целлюлозу, но на обычных средах, содержащих простые
сахара, они развиваются слабо, плохо переносят даже незначитель-
но повышенные концентрации Сахаров.
В рубце жвачных животных обитают специфичные облигат-
ные анаэробные целлюлозоразлагающие бактерии. Они вызывают
разложение целлюлозы кормов до глюкозы, которая затем сбражи-
вается с образованием органических кислот (уксусной, пропионо-
вой, масляной, молочной, муравьиной, янтарной и др.), спиртов и
газов (СО
2
и Н
2
). Разложение целлюлозы в рубце животных
осуществляется при участии кокковидных и палочковидных бакте-
рий: Ruminococcus flavefaciens, Ruminococcus albus, Bacteroides succino-
genes, Butyrivibrio fibrisolvens, Ruminobacter parvum. Бактерии рубца
очень важны для нормального питания жвачных животных.
Кратко остановимся на биохимической стороне процесса рас-
пада целлюлозы. Это линейный гомополисахарид, состоящий из
соединенных β-l,4-гликoзидными связями глюкозных остатков.
Молекулярная масса полисахарида — до 500 000. В молекуле целлю-
лозы может содержаться до 14 000 молекул β-D-глюкозы. Целлюлоз-
ные волокна представляют собой пучки микрофибрилл и мицеллы
(кристаллиты) — весьма плотно упакованные цепочки макромоле-
кул, чередующиеся с некристаллическими или паракристаллически-
ми участками. Последние в первую очередь подвергаются воздейст-
вию ферментов микроорганизмов.
Разложение целлюлозы с помощью микроорганизмов прохо-
дит в несколько этапов. Сначала идет ферментный гидролиз поли-
мера. Ферментная система, осуществляющая разложение целлюлозы
до глюкозы, носит название целлюлазного комплекса. В его состав
входят: эндо- β-1,4-глюканаза (1,4- β-D-глюкан-4-глюкогидролаза),
экзо-1,4- β-глюканаза (целлобиогидролаза), экзо-1,4- β-глюкозидаза
(1,4-D-глюкан-глюкогидролаза), целлобиаза ( β-глюкозидаза).
187
Процесс расщепления целлюлозы протекает следующим обра-
зом: при действии на целлюлозу фермента эндоглюконазы образу-
ются олигосахара различной степени полимеризации, а также цел-
лобиоза. Затем под влиянием эндоглюканазы и целлобиогидролазы
олигосахара гидролизуются до целлобиозы. Целлобиаза катализи-
рует расщепление целлобиозы на две молекулы глюкозы. Все фер-
менты целлюлазного комплекса бактерий, за исключением целлоби-
азы, внеклеточные. При аэробном разложении целлюлозы из обра-
зовавшейся глюкозы в основном получается два продукта — СО
2
и
Н
2
О. Могут накапливаться и небольшие количества органических
кислот.
В последние годы у ряда анаэробных целлюлозоразлагающих
бактерий были обнаружены на поверхности клеток высокомолеку-
лярные мультисубъединичные протуберантные структуры, ответст-
венные за разложение целлюлозы, получившие название целлюло-
сом. Это связанный с поверхностью клетки комплекс, содержащий
набор целлюлолитических ферментов, а также белки, не имеющие
ферментной активности. Входящие в комплекс ферменты представ-
лены эндоглюканазами, а также ферментами, играющими основную
роль в разложении целлюлозы. Комплекс обеспечивает прикрепле-
ние клеток к целлюлозе и ее высокоэффективный гидролиз.
При анаэробном распаде целлюлозы первоначальный продукт
ее гидролиза — глюкоза в дальнейшем подвергается сбраживанию, в
результате чего возникает много органических веществ, состав кото-
рых различается у отдельных культур микроорганизмов:
Мезофилы:
Clostridium omelianskii Этанол, уксусная, молочная
и муравьиная кислоты, СО
2
, Н
2
С. dissolvens Этанол, уксусная, молочная
и масляная кислоты, СО
2
, Н
2
С. cellobioparum Этанол, уксусная, муравьиная
и молочная кислоты, СО
2
, Н
2
Термофилы:
С. thermocellum Этанол, уксусная, молочная,
муравьиная кислоты, СО
2
, Н
2
Thermoanaerobacter ethanolicus Этанол
Раньше считали, что при брожении целлюлозы образуется ме-
тан СН
4
. Однако позднее было показано, что метан выделяется не
в результате деятельности бактерий, использующих полисахариды,
а вследствие жизнедеятельности вторичного бактериального насе-
ления — микробного сообщества, утилизирующего продукты разло-
жения целлюлозы. В конце этой пищевой цепи находятся метаноге-
ны (архебактерии), продуктом жизнедеятельности которых является
метан.
188
Имеющиеся в растениях моно- и дисахариды, а также низко-
молекулярные полисахариды (крахмал, инулин, камеди и т. п.) лег-
ко разрушаются различными микроорганизмами.
Разложение гемицеллюлозы. Гемицеллюлозы наряду с лиг-
нином и пектинами входят в состав межклеточного вещества расти-
тельных тканей, в значительных количествах содержатся в древеси-
не, соломе, кукурузных початках и т. д. Как и целлюлоза, они при-
сутствуют в клеточной стенке. Кроме растений, гемицеллюлозы
обнаружены у грибов и дрожжей, входят в состав внеклеточных по-
лисахаридов.
Гемицеллюлозы — гетерополисахариды, состоящие из пентоз
(ксилозы, арабинозы) или гексоз (глюкозы, маннозы, галактозы),
что отражено в их названиях: пентозаны (ксилан или арабан), ман-
наны, галактаны и т. д. Ксилан — полимер ксилозы — по содержа-
нию в растениях занимает второе место после целлюлозы. Солома и
луб содержат до 30% ксилана, древесина хвойных — 7—12, листвен-
ных пород — 20—25%. Молекулы ксилана состоят из остатков β-D-
ксилозы, соединенных 1,4-гликозидными связями. Многие геми-
целлюлозы содержат также уроновые кислоты.
Перечисленные вещества активно разлагаются грибами,
аэробными и анаэробными бактериями. В этих процессах участвует
значительно больше видов микроорганизмов, чем в разложении
целлюлозы. Многие из них не очень специфичны и кроме полиса-
харидов способны усваивать органические кислоты и многие прос-
тые сахара.
Микробное население, участвующее в разложении гемицел-
люлоз растений, очень разнообразно. Это связано с неодинаковым
химическим составом указанных полисахаридов у разных растений,
что оказывает влияние и на характер конечных продуктов распада.
К микроорганизмам, обладающим гемицеллюлозоразлагающими
свойствами, относят бактерии родов Clostridium, Bacillus, Cytophaga,
Sporocytophaga (например, Sporocytophaga myxococcoides), Vibrio, Strep-
tomyces; грибы родов Aspergillus, Rhizopus, Forties, Polyporus и др. Геми-
целлюлозы в кислых почвах обычно разлагают грибы, в нейтральных
и щелочных — бациллы, Sporocytophaga и ряд других бактерий.
Большая молекулярная масса гемицеллюлоз не позволяет их мо-
лекулам проникать через цитоплазматическую мембрану бактериаль-
ной клетки. Молекулы должны быть превращены в простые соедине-
ния сахара, и только тогда микроорганизмы смогут их использовать.
Ферменты, катализующие расщепление гемицеллюлоз, носят
название гемицеллюлаз (ксиланаз). Многие микроорганизмы, обла-
дающие способностью к синтезу целлюлазного комплекса, имеют
и ксиланазу.
Первое время, после того как растительные остатки попадают
в почву, гемицеллюлозная фракция полисахаридов разлагается со
значительной скоростью, затем процесс замедляется. Это, вероятно,
189

результат химической гетерогенности гемицеллюлозных фракций:
одни разлагаются медленнее, другие — быстрее. Скорость превра-
щения гемицеллюлоз микроорганизмами почвы зависит и от усло-
вий среды — температуры, влажности, рН среды и т. д.
Разложение лигнина. Растения, особенно древесные, содер-
жат большое количество лигнина во вторичных слоях клеточной
оболочки и как основной компонент в составе межклеточного ве-
щества и вторичных слоях клеточной стенки. В молодых растениях
количество лигнина относительно невелико, но с возрастом его со-
держание в тканях увеличивается. Молодые травы содержат от 3 до
6% лигнина (на сухое вещество), древесина разных деревьев — от 18
до 30%. Вероятно, это соединение никогда не встречается в свобод-
ном виде, обычно оно связано с полисахаридами.
Лигнин, содержащийся в растениях разных видов, родов и се-
мейств растительного царства, химически неоднороден. Даже в од-
ном растении в зависимости от фазы развития химический состав
лигнина может меняться.
Молекулярная масса лигнина 1000—10 000, он нерастворим в
воде и в большинстве органических растворителей. Молекула лиг-
нина содержит только три элемента — углерод, водород и кислород,
однако это весьма сложное соединение, состоящее из большого
числа полимеризованных мономерных блоков, которые представля-
ют собой производные фенилпропана. Главный мономер лигнина —
конифериловый спирт, он составляет скелет лигнина хвойных по-
род. Лигнин лиственных пород состоит из кониферилового и сина-
пового спиртов, лигнин злаковых растений имеет еще и кумаровый
спирт.
Лигнин устойчив к воздействию микроорганизмов, разлагается
значительно медленнее, чем целлюлоза и гемицеллюлоза. В аэроб-
ном разложении лигнина могут принимать участие многие предста-
вители класса Basidiomycetes. Так, при умеренной температуре лиг-
нин усваивают многие высшие грибы родов Clavaria, Armillariella,
Fomes, Polystictus, Polyporus и Ustilina. Активны по отношению к лиг-
нину Fusarium lactis, F. nivala, Trichoderma lignorum, Alternaria tenuis,
Stremphylium botryosum.
В почве есть аэробные бактерии рода Pseudomonas, участвую-
щие в термофильном разложении лигнина. Бактерии рода Clostridi-
um разлагают это соединение в анаэробных условиях. Считают, что
лигнин может трансформироваться и актиномицетами.
Лигнин деполимеризуется до простых ароматических веществ,
таких, как ванилин и другие метоксилированные ароматические
структуры. Ферментная система микроорганизмов, воздействую-
щих на лигнин, внеклеточная и представлена лигниназами — спе-
цифичными пероксидазами. В связи с тем что лигнин разрушается
190
относительно медленно, он накапливается в почве, и продукты раз-
ложения служат основой при образовании гумусовых веществ.
Разложение пектиновых веществ. Межклеточные вещества
растительных тканей — пектины — найдены в так называемых сре-
динных пластинках, расположенных между отдельными клетками
тканей растений. Первичные и вторичные клеточные стенки также
содержат полисахариды пектинового типа. Пектиновые вещества —
сложные полисахариды, полигалактурониды, это неразветвленные
полимеры, состоящие из остатков D-галактуроновых кислот, соеди-
ненных 1,4-гликозидными связями.
Существуют три типа пектиновых веществ: протопектин —
водонерастворимая составная часть клеточной стенки; пектин — во-
дорастворимый полимер галактуроновой кислоты, содержащей метил-
эфирные связи; пектиновая кислота — водорастворимый полимер
галактуроновой кислоты, свободный от метилэфирных связей. Пек-
тиновая кислота образована длинными цепочками галактуроновых
кислот, способных после обработки кальциевыми солями к форми-
рованию твердого пектинового геля.
Бактерии и грибы воздействуют на пектин, протопектин и пек-
тиновую кислоту в аэробных и анаэробных условиях. В почве обна-
ружено большое число микроорганизмов, разлагающих пектиновые
вещества (до 1 млн клеток на 1 г почвы). Высокой пектинолитиче-
ской активностью обладают представители семейства Ваcillасеае —
аэробные бактерии рода Bacillus (В. macerans, В. polymyxa) и ана-
эробные рода Clostridium (С. pectinovorum, С. felsineum, С. aurantibu-
tyricum, С. pectinolyticum, С. соralliпит, С. flavum и др.), а также мно-
гие грибы. Пектины разлагаются и под влиянием фитопатогенных
грибов (Botrytis cinerea, Fusarium oxysporum f. lycopersici) и бактерий
Erwinia carotovora), использующих эту свою способность для про-
никновения в ткани растений.
Микроорганизмы синтезируют следующие экзоферменты (эс-
теразы и деполимеразы), катализующие распад пектиновых веществ:
• протопектиназу, участвующую в разложении протопектина
с образованием растворимого пектина;
• пектинэстеразу, гидролизующую метилэфирную связь пек-
тина с образованием пектиновой кислоты и метилового
спирта;
• пектиназу (полигалактуроназу), разрушающую связи между
отдельными составляющими галактуроновой кислоты, пек-
тина или пектиновой кислоты с образованием небольших
цепочек и в конечном счете свободной D-галактуроновой
кислоты.
При гидролизе пектиновой кислоты пектиназой на первых
этапах аккумулируются только небольшие количества свободной
D-галактуроновой кислоты. Обычно при участии ферментов разла-
191

гаются ди-, три-, тетра- и пентагалактуроновые кислоты. В после-
дующих этапах гидролиза длинные молекулы распадаются под влия-
нием каталитической деятельности пектиназы и накапливаются
свободная D-галактуроновая кислота и другие соединения.
Распад пектиновой кислоты может быть выражен следующей
схемой:
Продукты распада пектиновой кислоты (галактоза, арабиноза
и др.) подвергаются окислению или сбраживанию при воздействии
разнообразных микроорганизмов. При анаэробиозе это маслянокис-
лые бактерии, относящиеся к роду Clostridium (С. pectinovorum,
С. felsineum и др., рис. 40). Продуктами распада С. pectinovorum слу-
жат масляная и уксусная кислоты, а также газы Н
2
и СО
2
, а в куль-
туре С. felsineum кроме указанных веществ образуется и небольшое
количество ацетона и бутанола.
Разложение пектиновых веществ наблюдается при мочке лу-
боволокнистых растений — льна, конопли, кенафа, джута, канатни-
ка и др. Целлюлозные волокна указанных растений склеены с окру-
жающими их тканями пектином. Для отделения волокон необходи-
мо разложение пектина, для чего используют пектинолитическое
действие ферментов анаэробных бактерий.
При водной мочке после погружения в воду стебли льна набу-
хают. При этом экстрагируются водорастворимые вещества (сахара,
глюкозиды, таннины, растворимые соединения азота и пигменты) и
начинают развиваться бактерии. Сначала размножаются аэробные
формы, так как вода содержит кислород и питательные вещества,
способствующие их развитию. Дрожжи и плесневые грибы развива-
ются на поверхности среды. По-
глощение аэробными микроорга-
низмами из жидкости кислорода
обусловливает создание в среде
анаэробных условий. Начинают раз-
виваться факультативно анаэроб-
ные бесспоровые бактерии, близ-
кие к Escherichia coli. Постепенно
в среде накапливаются органиче-
ские кислоты (в том числе молоч-
ная) и выделяются газы СО
2
и Н
2
.
Отделение волокон происхо-
дит во время основной стадии бро-
жения, когда анаэробные бактерии
192
тиna С. pectinovorum начинают расщеплять пектин. Накопление ор-
ганических кислот приводит к прекращению деятельности С. pecti-
novorum. Последнего сменяет более кислотоустойчивый микроорга-
низм С. felsineum. В результате воздействия ферментов микроорга-
низмов двух указанных видов на пектиновые вещества паренхимной
ткани от коры и древесины отделяются пучки волокон.
На льнозаводах тепловую мочку льноволокна выполняют в
особых чанах — мочилах при 32—38 °С в течение трех—пяти суток.
Для ускорения мочки и увеличения выхода длинного волокна пред-
ложен препарат пектолитин, содержащий споры активного пектин-
разлагающего микроорганизма — С. felsineum. Внесение пектолити-
на в мочильную жидкость приводит к ускорению процесса в сред-
нем на 27%, повышается и выход длинного волокна, его качество.
В производство льняного волокна внедряют также препараты пекто-
литических ферментов.
Применяют и аэробную мочку льна и других лубоволокнис-
тых культур. В этом случае пектиновые вещества разрушаются
аэробными микроорганизмами. Предварительно пектиновые веще-
ства гидролизуются до галактуроновых кислот, галактозы, арабино-
чы, ксилозы, уксусной кислоты и метанола, а затем начинается
окисление (бактериями, дрожжами, грибами) возникших соедине-
ний до СО
2
и Н
2
О.
Существует несколько способов аэробной мочки льна. Рас-
стил, или росяная мочка, — самый старый и наиболее примитивный
биологический способ получения волокна. При этом лен в осеннее
время года расстилают на траве. Широкий доступ воздуха, система-
тичное и порой длительное отсутствие влаги, воздействие света и
атмосферных осадков, суточные колебания температуры обусловли-
вают длительность процесса (три—восемь недель) и преобладание в
нем не бактерий, а плесневых грибов. Насчитывают до 80 видов
грибов, участвующих в росяной мочке. Первое время после рассти-
ла стеблей на траве наблюдается развитие бактерий, а затем преоб-
ладают грибы — Rhizopus, Aureobasidium, Alternaria и Gonatobotrys.
Развитие перечисленных представителей грибов указывает на опти-
мальную степень мочки льносоломы. Видовой состав микрофлоры
при мочке определяется географическими и почвенно-климатиче-
скими условиями. При мочке расстилом существует опасность, что
многие грибы (например, Cladosporium herbarum, Trichoderma lig-
norum), появляющиеся на льносоломе после Rhizopus, Alternaria и
других, разрушат целлюлозу, т. е. волокно. Особенно часто это про-
исходит при перележивании разостланной соломы.
В благоприятных атмосферных условиях (теплая и влажная
погода), сочетающихся с тщательным уходом за соломой (защита от
спутывания, переворачивание рядков, своевременная уборка со
стлища), расстил дает вполне удовлетворительное по качеству и вы-
ходу волокно.
7 Микробиология 193
Контрольные вопросы и задания
1. Какие микроорганизмы служат возбудителями молочнокислого брожения?
2. В чем сущность пропионовокислого брожения? 3. Чем отличается окисле-
ние углеводов при участии микроорганизмов от различного типа брожений?
Глава 10 Превращение микроорганизмами
соединений азота
От азотного питания растений во многом зависит величина
урожая сельскохозяйственных культур. Большинству растений не-
доступен газообразный азот, в огромном количестве находящийся
в воздухе, а из разнообразных азотных соединений, встречающихся
в почве, они могут усваивать только минеральные. Поэтому столь
важен вопрос о превращениях соединений азота в почве под воздей-
ствием микроорганизмов.
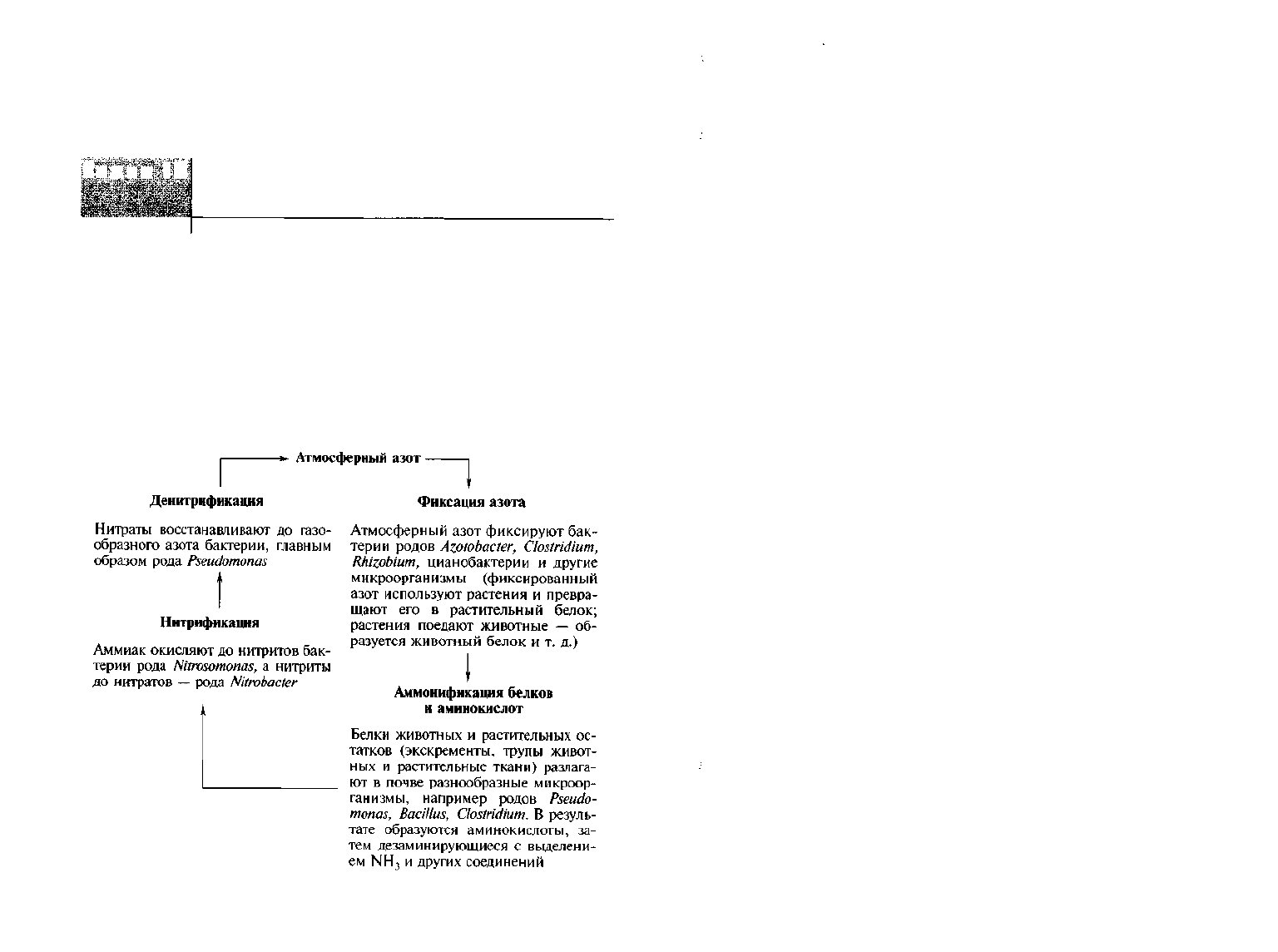
Превращения азота и содержащих этот элемент соединений в
почве довольно сложны, но в них можно выделить основные на-
правления, определяющие круговорот азота в природе:
194
Некоторую часть атмосферного азота связывают свободножи-
вущие или находящиеся в симбиозе с растениями микроорганизмы.
Данный процесс обогащает азотом и почву, и растения. Органиче-
ские азотсодержащие соединения в тканях растений и животных,
попадая в почву, подвергаются минерализации до аммонийных со-
единений. Часть растительных остатков трансформируется в темно-
окрашенное, содержащее азот вещество, — гумус.
Аммонийная форма азота подвергается в почве окислению
нитрифицирующими бактериями с образованием солей азотной
кислоты. При определенных условиях нитраты могут восстанавли-
ваться до молекулярного азота и улетучиваются из почвы. Значи-
тельное количество азотсодержащих соединений микроорганизмы
ассимилируют, а азот в органических формах практически недосту-
пен растениям.
Приведенные примеры показывают, что микроорганизмы
способны вызывать как мобилизационные процессы и накапливать
доступные для растений минеральные азотсодержащие вещества,
гак и диаметрально противоположные им — иммобилизационные,
обедняющие почву ценными для растений соединениями.
10.1. Минерализация азота
Среди органических соединений, составляющих клетку, первое мес-
то по количеству занимают белки — на их долю приходится не ме-
нее 50% сухой массы клетки. Значительная часть белков попадает
в почву с остатками отмерших растений, животных и микроорга-
низмов. При разложении белков и других азотсодержащих соедине-
ний в почве при участии микроорганизмов азот освобождается в ви-
де аммиака. Указанный процесс называют аммонификацией, или ми-
нерализацией азота.
Белки подвергаются разложению как аэробными, так и анаэ-
робными бактериями, а также актиномицетами и грибами. Осо-
бенно активны представители семейства Pseudomonadaceae, рода
Pseudomonas (P. fluorescens, P. aeruginosa), семейства ВасШасеае, рода
Bacillus (В. mycoides, В. cereus, В. subtilis) и рода Clostridium (С. spo-
rogenes, С. putrificus) (рис. 41), семейства Enterobacteriaceae, рода Pro-
teus (P. vulgaris) и др.
В состав белков обычно входит 20 ос-аминокислот. Аминокис-
юты в полимерной цепи белка располагаются так, что конец одной
аминокислоты связан с началом другой пептидной связью. Такие
полимерные молекулы, называемые полипептидными цепями, со-
держат до сотен аминокислотных звеньев. Белковая молекула вклю-
чает одну или несколько полипептидных цепей. По составу белки
подразделяют на простые и сложные. При гидролизе простых бел-
195

Рис. 41. Аммонифицирующие бактерии:
А — Bacillus sp.; Б — Bacillus cereus; В — Clostridium sp. (по: С. Робиноу)
ков образуются только аминокислоты, сложных — как аминокисло-
ты, так и другие органические и неорганические продукты. Небел-
ковая (не содержащая аминокислот) часть молекулы сложного белка —
это его простетическая группа. В зависимости от ее состава слож-
ные белки называют нуклеопротеидами, липопротеидами, металло-
протеидами и гликопротеидами.
Молекулы белков и большинства пептидов велики и не могут
проходить через цитоплазматическую мембрану микроорганизмов.
Поэтому они расщепляются экзоферментами. Протеолитические
ферменты, или протеазы, выделяемые клетками микроорганизмов в
окружающую среду, осуществляют гидролиз ряда пептидных связей
в молекулах белков. Образующиеся при этом частицы белковой мо-
лекулы (полипептиды и олигопептиды) поступают внутрь клеток
микроорганизмов, где разрушаются внутриклеточными протеолити-
196
ческими ферментами — пептидазами до свободных аминокислот.
Последние используются для синтеза белков клетки или подверга-
ются дальнейшему расщеплению.
Внутриклеточное или внеклеточное расщепление аминокис-
лот может идти следующими четырьмя путями:
Аминокислоты минерализуются с различной скоростью. Не-
которые из них (треонин, метионин) более устойчивы, другие (ар-
гинин, триптофан), наоборот, разлагаются легко. После дезами-
нирования углеродный остаток подвергается воздействию микробов
к аэробных или анаэробных условиях до образования СО, и различ-
ных органических соединений. Если в среде есть амиды, то перво-
начально они разлагаются до аминокислот и только затем уже могут
быть трансформированы тем или иным путем. Например, аспарагин
под действием фермента аспарагиназы превращается в аспарагино-
вую кислоту:
При аэробном распаде белка основными конечными продук-
тами процесса бывают СО
2
, аммиак, сульфаты и вода. В анаэробных
условиях при распаде белка образуются аммиак, амины, СО
2
, орга-
нические кислоты (жирные и ароматические — бензойная, ферули-
новая и др.), меркаптаны, а также вещества с неприятным запахом —
индол, скатол и сероводород.
При анаэробном разрушении белков могут образовываться
токсичные соединения, в частности первичные амины (диамины)
или птомаины. К числу последних относят кадаверин, который по-
лучается из лизина:
Накапливающиеся в анаэробных условиях в почве продукты
разложения белков фитотоксичны, поэтому они нередко угнетающе
действуют на растения и снижают урожайность.
При разрушении сложных белков сначала освобождаются ос-
новные составляющие — белок и связанная с ним простатическая
197
группа. В дальнейшем эти соединения (каждое самостоятельно) под-
вергаются более глубокой трансформации.
Разложение нуклеиновых кислот. Нуклеиновые кислоты вхо-
дят в состав нуклеопротеидов. Последних особенно много в клеточ-
ных ядрах. Нуклеиновые кислоты — РНК и ДНК — органические
вещества большой молекулярной массы, представляющие собой по-
лимеры. При их гидролизе освобождаются пуриновые и пиримиди-
новые соединения, сахар и фосфорная кислота. Сахар в РНК пред-
ставлен рибозой, в ДНК — дезоксирибозой. Пурины — аденин и гу-
анин найдены как в молекулах РНК, так и в молекулах ДНК. Из
пиримидинов — цитозин обнаружен в РНК и ДНК, урацил только в
РНК, а тимин — только в ДНК.
Длинные молекулы нуклеиновых кислот при разложении де-
полимеризуются. Сначала отщепляются небольшие фрагменты, ко-
торые затем распадаются на отдельные мононуклеотиды. Процесс
расщепления идет при участии ферментов рибонуклеазы и дезокси-
рибонуклеазы, синтезируемых некоторыми видами грибов, актино-
мицетами и рядом бактерий. На следующем этапе от мононуклеоти-
дов под воздействием нуклеотидаз отщепляется фосфорная кислота,
затем — сахар, пуриновые и пиримидиновые основания.
В зависимости от типа обмена веществ микроорганизмов сахар
в дальнейшем может окисляться кислородом до СО
2
и Н
2
О или под-
вергаться брожению с образованием органических кислот и спир-
тов. Азотсодержащие основания разлагаются до мочевины и амино-
кислот и в конце концов до аммиака и органических кислот.
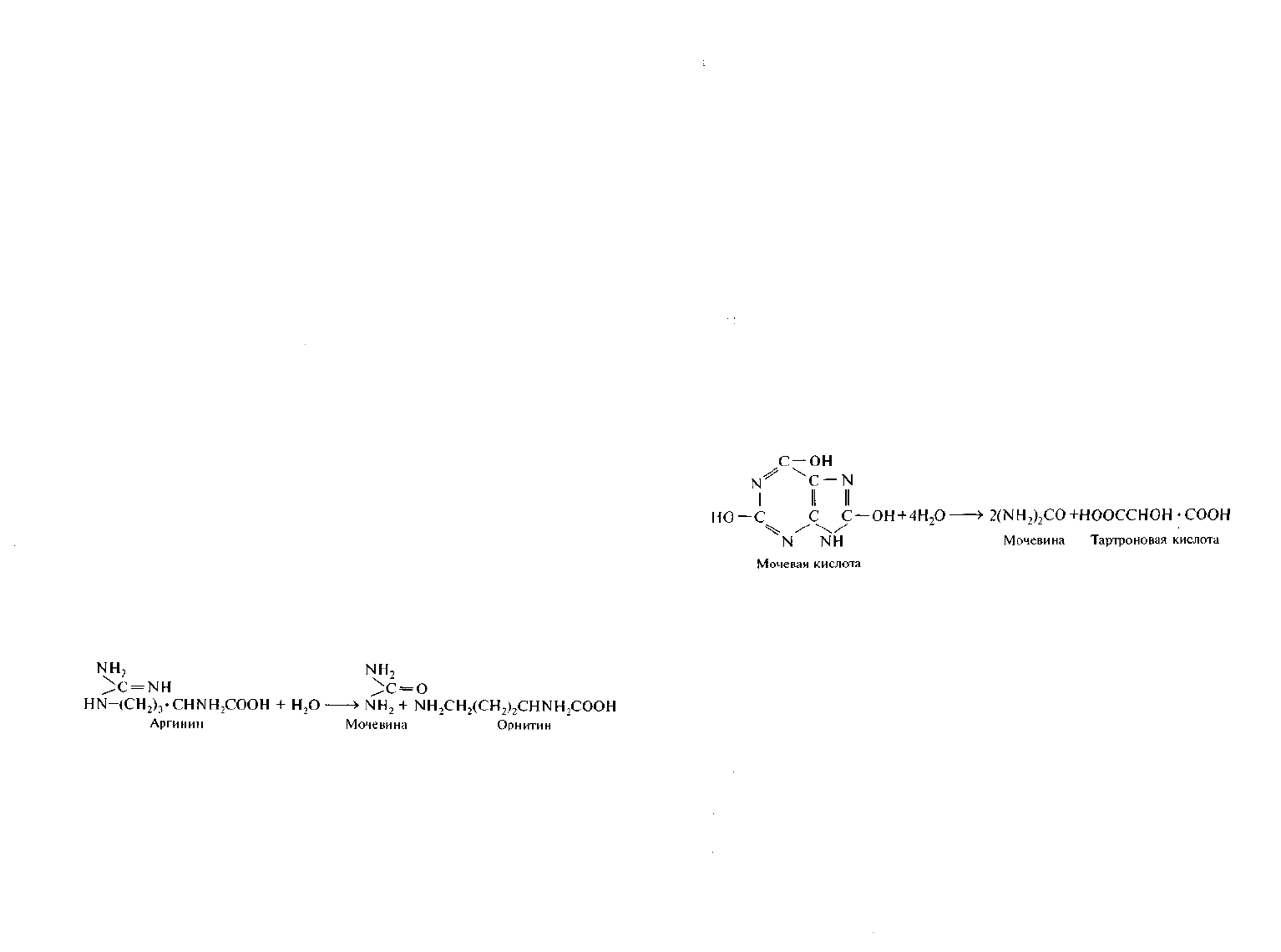
Разложение мочевины. К азотсодержащим органическим со-
единениям, часто встречающимся в природе, относятся мочевина,
мочевая и гиппуровая кислоты. Мочевина присутствует в моче че-
ловека и животных, ее могут синтезировать почвенные грибы. Так,
до 13% сухой массы плодовых тел шампиньонов составляет мочеви-
на. Это же соединение образуется при гидролитическом распаде ар-
гинина под действием фермента аргиназы:
На земном шаре за год организмы синтезируют до 30 млн т
мочевины, что составляет существенные ресурсы азота: мочевина
содержит 46% этого элемента и используется как удобрение. Под
действием микроорганизмов, содержащих фермент уреазу, мочевина •
в несколько этапов превращается в аммиак и диоксид углерода:
CO(NH
2
)
2
+ 2Н
2
О -----> (NH
4
)
2
CO
3
Мочевина
198
Образующаяся на первом этапе углеаммиачная соль малоус-
ойчива и быстро разлагается:
(NH
4
)
2
CO
3
= 2NH
3
+ СО
2
+ Н
2
О
Многие бактерии и грибы, синтезирующие уреазу, могут ис-
пользовать мочевину как источник азота для синтеза белков. Обычно
бактерии, разлагающие мочевину, называют уробактериями. Эти орга-
низмы могут развиваться при высокой щелочности среды (рН 9—10),
что позволяет им вызывать распад значительного количества моче-
вины до аммиака. Из специфических уробактерий наиболее важны:
Micrococcus urea из семейства Micrococcaceae, Bacillus pasteurii из се-
мейства Bacillaceae, а также Sporosarcina urea.
Физиологический смысл распада мочевины, по-видимому,
сводится к переводу аминной формы азота в более легкоусвояемую
аммиачную.
Разложение мочевой и гиппуровой кислот. Мочевая и гиппу-
ровая кислоты также играют важную роль в белковом обмене мле-
копитающих, пресмыкающихся, насекомых и птиц. В экскрементах
змей содержится до 90% мочевой кислоты, а в помете птиц — 25%.
В моче млекопитающих содержание мочевой кислоты незначитель-
но. Мочевая и гиппуровая кислоты быстро распадаются под влия-
нием гидролитических ферментов ряда микроорганизмов:
Разложение мочевой кислоты в местах скопления помета
птиц (гуано) в условиях засушливого климата приводит к накоп-
лению нитратов. Образующийся в таких местах в массе аммиак под-
вергается окислению нитрифицирующими бактериями. В связи
с низкой влажностью нитраты из гуано не вымываются, а накапли-
ваются. Все это обусловило возникновение богатых залежей нитра-
тов в Чили, Перу, Южной Африке и на островах Карибского моря.
Гуано содержит 9% азота, 13% фосфорной кислоты, калий, каль-
ций, различные микроэлементы, поэтому используется как удобре-
ние в сельском хозяйстве.
Разложение цианамида кальция. Цианамид кальция (CaCN
2
)
используют как азотное удобрение, которое растения самостоя-
тельно не ассимилируют, но в почве оно быстро превращается в ам-
миак. Разложение цианамида кальция проходит в три этапа. Первый
ротекает самопроизвольно под влиянием почвенной влаги до обра-
ования цианамида. Ряд почвенных катионов (Са, Mn, Fe и т. д.)
199

вызывает превращение цианамида в мочевину. Далее гидролиз мо-
чевины происходит под влиянием уробактерий:
CaCN
2
+ 2Н
2
О > H
2
CN
2
+ Са(ОН)
2
H
2
CN
2
+ Н
2
О > CO(NH
2
)
2
CO(NH
2
)
2
+ Н
2
О > 2NH
3
+ СО
2
Разложение хитина. Разложение хитина осуществляют мно-
гие почвенные микроорганизмы, он постоянно присутствует в поч-
ве. Это азотсодержащий полисахарид, полимер ацетилглюкозамина.
Хитин содержится в наружном скелете беспозвоночных животных,
в панцирных покровах насекомых, в клеточной стенке многих гри-
бов, в частности базидиомицетов и аскомицетов.
Способностью разлагать хитин обладают около 50 видов бак-
терий родов Pseudomonas, Flavobacterium, Bacillus, Cytophaga, Strepto-
myces, Nocardia и Micromonospora. Из грибов активно воздействуют
на это вещество мукоровые грибы и аспергиллы (Aspergillus fumiga-
tus), а также Mortierella. Особенно легко разлагается хитин актино-
мицетами. Известны и миксобактерии, усваивающие хитин, напри-
мер скользящая бактерия Chitinophaga pinensis.
Под действием синтезируемого микроорганизмами фермента
хитиназы хитин вначале разлагается на хитобиозу и хитотриозу, по-
путно образуется небольшое количество N-ацетилглюкозамина.
Затем хитобиоза и хитотриоза расщепляются в присутствии хитоби-
азы до уксусной кислоты, глюкозы и аммиака.
10.2. Нитрификация
Аммиак, образующийся в почве, навозе и воде при разложении ор-
ганических веществ, довольно быстро окисляется до азотистой, а за-
тем азотной кислоты. Такой процесс называют нитрификацией.
До середины XIX в., точнее, до работ Л. Пастера явление об-
разования нитратов объясняли как химическую реакцию окисления
аммиака атмосферным кислородом, причем предполагалось, что поч-
ва в этом процессе играет роль катализатора. Л. Пастер предположил,
что образование нитратов — микробиологический процесс. Первые
экспериментальные доказательства его гипотезы были получены
Т. Шлезингом и А. Мюнцем в 1879 г. Исследователи пропускали
сточные воды через длинную колонку с песком и СаСО
3
. При фильт-
рации аммиак постепенно исчезал и появлялись нитраты. Нагревание
колонки или внесение антисептиков прекращало окисление аммиака.
Однако выделить культуры возбудителей нитрификации не
удалось ни упомянутым исследователям, ни микробиологам, продол-
жавшим изучение нитрификации. Лишь в 1890—1892 гг. С. Н. Вино-
градский, применив особую методику, изолировал чистые культуры
нитрификаторов. Ученый предположил, что нитрифицирующие бак-
200
терии не растут на обычных питательных средах, содержащих орга-
нические вещества, это объяснило неудачи его предшественников.
Действительно, нитрификаторы оказались хемолитоавтотро-
фами, т. е. бактериями, использующими энергию окисления амми-
ака или азотистой кислоты для синтеза органических веществ из
СО
2
(хемосинтез). Поэтому их клетки очень чувствительны к при-
сутствию в среде органических соединений. Нитрифицирующие
бактерии удалось выделить на минеральных питательных средах.
С. Н. Виноградский установил, что существуют две группы ни-
трификаторов: одна осуществляет окисление аммиака до азотистой
кислоты (NH
+
4
> NO
-
2
) — первая фаза нитрификации, дру-
гая — окисление азотистой кислоты до азотной (NO
-
2
> NO
-
3
) —
вторая фаза нитрификации.
Представителей обеих групп относят к семейству Nitrobacte-
riaceae. Это одноклеточные грамотрицательные бактерии. Среди ни-
трифицирующих бактерий есть палочковидные клетки, эллиптиче-
ские, сферические, извитые и дольчатые, плеоморфные. Размеры
клеток колеблются от 0,3 до 1 мкм в ширину и от 1 до 3 мкм в дли-
ну. Существуют подвижные и неподвижные формы с полярным,
субполярным и перитрихальным жгутикованием.
Размножаются бактерии-нитрификаторы в основном делением,
зa исключением Nitrobacter, для которого характерно почкование. По-
чти у всех нитрификаторов хорошо развита система внутрицитоплаз-
матических мембран, значительно различающихся по форме и распо-
ложению в клетках отдельных видов. Мембраны цитоплазмы подоб-
ны мембранам фотосинтезирующих пурпурных бактерий.
Бактерии первой фазы нитрификации представлены ро-
дами: Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosolobus и Nitroso-
vibrio. Наиболее детально к настоящему времени изучен Nitrosomonas
europaea (рис. 42, А). Он представляет собой короткие овальные па-
лочки размером 0,8—1 х 1—2 мкм. В жидкой культуре клетки Ni-
trosomonas проходят ряд стадий развития. Две основные из них пред-
ставлены подвижной формой и неподвижными зооглеями. Подвиж-
ная форма обладает субполярным жгутиком или пучком жгутиков.
Описаны представители и других родов бактерий, вызываю-
щих первую фазу нитрификации.
Вторую фазу нитрификации осуществляют представители
родов Nitrobacter, Nitrospira и Nitrococcus. Наибольшее число ис-
следований проведено с Nitrobacter winogradskyi (рис. 42, Б), однако
описаны и другие виды (например, Nitrobacter agilis). Клетки нитро-
бактepa имеют удлиненную, клиновидную или грушевидную фор-
му, более узкий конец часто загнут в клювик, размеры клеток —
0,6—0,8 х 1—2 мкм. При почковании дочерняя клетка обычно под-
вижна, так как имеет один полярный жгутик. Известно чередование
в цикле развития подвижной и неподвижной стадий.
201

Рис. 42. Нитрифицирующие бактерии:
А — Nitrosomonas europaea; Б — Nitrobacter winogradskyi
Описаны и другие виды бактерий, вызывающие вторую фазу
нитрификации.
Нитрифицирующие бактерии культивируют на простых мине-
ральных средах, содержащих аммиак или нитриты (окисляемые суб-
страты) и диоксид углерода (основной источник углерода). Источником
азота для этих организмов служат аммиак, гидроксиламин и нитриты.
Нитрифицирующие бактерии развиваются при рН 6,0—
8,6, оптимум реакции среды составляет рН 7,5—8,0. При значениях
ниже рН 6 и выше рН 9,2 бактерии не развиваются. Оптимальная
температура для развития нитрификаторов 25—30 °С. Изучение от-
ношения различных штаммов Nitrosomonas europaea к температуре
показало, что некоторые из них имеют оптимум развития при 26 °С
или около 40 °С, другие способны довольно быстро расти при 4 °С.
Нитрификаторы — облигатные аэробы
1
. Используя кислород
воздуха, они окисляют аммиак до азотистой кислоты (первая фаза
нитрификации):
NH
+
4
+ 3/
2
О
2
> NO
2
-
+ Н
2
О + 2H
+
1
В последние годы обнаружена способность бактерий к анаэробному
окислению аммиака. Этот процесс, получивший название «анаммокс» (Ап-
аттох), играет важную роль при очистке сточных иод. Осуществляющие его
бактерии относятся к группе планктомицетов. (Прим. ред.)
202
а затем азотистую кислоту до азотной (вторая фаза нитрификации):
NO
2
-
+ 1/
2
О
2
> NO
3
-
Следовательно, аммиак — продукт жизнедеятельности ам-
монифицирующих бактерий — использует для получения энергии
Nitrosomonas, а нитриты, образующиеся в процессе жизнедеятель-
ности последних, служат источником энергии для Nitrobacter.
Согласно современным представлениям, процесс нитрифи-
кации осуществляется на цитоплазматической и внутрицитоплазма-
тических мембранах и проходит в несколько этапов. Первым про-
дуктом окисления аммиака становится гидроксиламин, затем пре-
вращающийся в нитроксил (NOH) или пероксонитрит (ONOOH),
последний, в свою очередь, преобразуется в дальнейшем в нитрит,
а нитрит в нитрат. Весь процесс нитрификации иллюстрирует сле-
дующая схема:
Нитроксил, как и гидроксиламин, по-видимому, может диме-
ризоваться в гипонитрит или превращаться в закись азота N
2
O —
побочный продукт нитрификации. Кроме первой реакции (образо-
вания гидроксиламина из аммония), все последующие превращения
сопровождаются синтезом макроэргических связей в виде АТФ.
Нитрификаторы осуществляют фиксацию СО
2
через восста-
новительный пентозофосфатный цикл (цикл Кальвина). В результа-
те последующих реакций образуются не только углеводы, но и дру-
гие важные для бактерий соединения — белки, нуклеиновые кисло-
ты, жиры и т. д.
Долгое время нитрифицирующих бактерий относили к облигат-
ным хемолитоавтотрофам. Позднее были получены данные о спо-
собности этих бактерий использовать некоторые органические ве-
щества. Так, отмечено стимулирующее действие на рост Nitrobacter
нитрита, дрожжевого автолизата, пиридоксина, глутаминовой кис-
лоты и серина. Предполагают, что некоторые нитрифицирующие
бактерии обладают способностью переключаться с автотрофного на
гетеротрофное питание. Однако нитрификаторы не растут на обыч-
ных питательных средах, так как большое количество легкоусвояе-
мых органических веществ, содержащихся в таких средах, задержи-
вает их развитие. Однако в природе такие бактерии хорошо развива-
ются в черноземах, навозе, компостах, т. е. в местах, где содержится
много органического вещества.
203