Дуус П. Топический диагноз в неврологии
Подождите немного. Документ загружается.


108
3 Ствол мозга
ной борозды), а оттуда по затылочно5
тектальному пути — к ядрам нервов,
иннервирующих мышцы глазных яб5
лок. В результате благодаря оптокине$
тическому процессу изображение вновь
начинает проецироваться на область
желтых пятен. При этом глазные ябло5
ки совершают толчкообразные движе5
ния (оптокинетический Та5
кой нистагм возникает при выглядыва5
нии из окна движущегося поезда, или
чтении, или, в эксперименте, при кон5
центрировании внимания на медленно
поворачивающемся цилиндре, окра5
шенном чередующимися черными и бе5
лыми полосами. Толчкообразные дви5
жения глаз возникают в направлении,
противоположном движению взгляда.
Оптокинетический нистагм исчезает
при перерыве вышеописанной рефлек5
торной дуги на любом уровне.
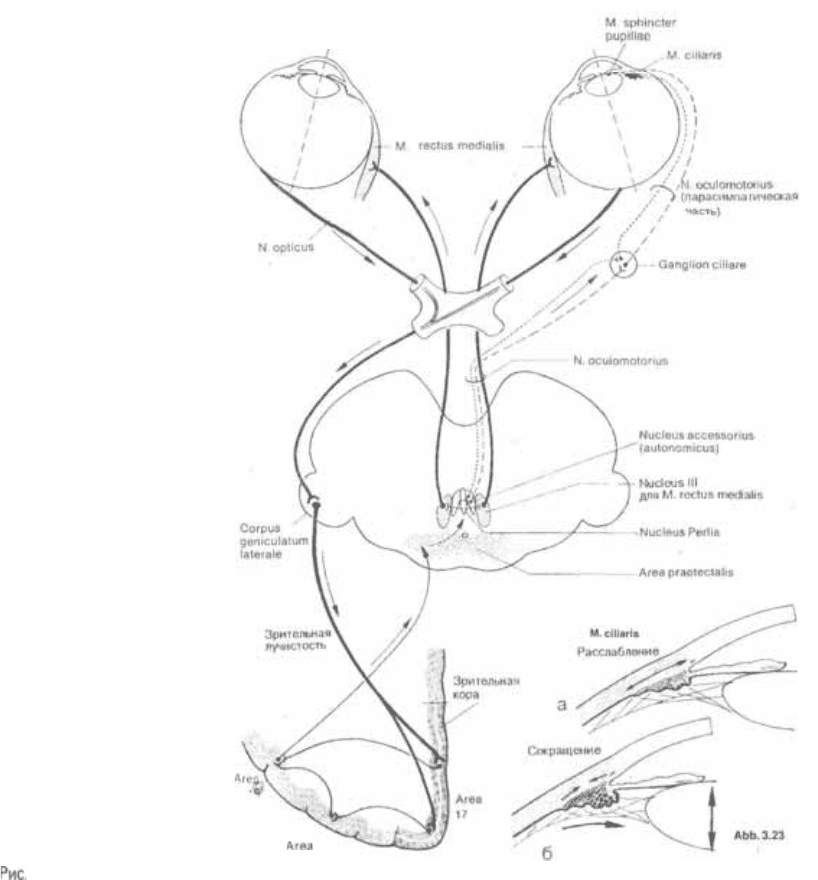
Конвергенция и аккомодация
Приближение предмета, находящегося
в центре поля зрения, вызывает раз5
личные рефлексы. Это рефлексы кон5
вергенции и (рис. 3.23).
При этом происходит следующее:
1. Конвергенция: обе внутренние прямые
мышцы начинают
синхронно, поэтому оси обоих глаз5
ных яблок поворачиваются по на5
правлению к предмету. В результате
этого изображение предмета проеци5
руется точно на соответствующие об5
ласти сетчаток, или на зоны наилуч5
шего видения.
2. Аккомодация: благодаря сокращению
цилиарной (ресничной) мышцы хру5
сталики расслабляются и становят5
ся более выпуклыми. Вследствие это5
го изображение приближающегося
предмета фокусируется на сетчатку.
(При взгляде вдаль расслабление ци5
лиарной мышцы приводит к упло5
щению хрусталика).
3. Сужение зрачка. Сужение зрачка обес5
печивает сохранение четких контуров
изображения предмета на сетчатке,
наподобие тому, как сужение диаф5
рагмы в фотоаппарате увеличивает
четкость изображения.
Все эти три реакции могут быть вы5
званы произвольной фиксацией на нахо5
дящемся рядом предмете. Они же воз5
никают рефлекторно при внезапном
приближении удаленного предмета. Аф5
ферентные импульсы идут от сетчатки
к коре шпорной области. Оттуда эффе5
рентные импульсы направляются через
претектальную область к парасимпати5
ческим ядрам Перлиа, расположенным
по средней линии вентральнее ядер
Эдингера5Вестфаля. Импульсы от этой
группы ядер идут к
нейронам, иннер5
вирующим обе внутренние прямые
мышцы (для конвергенции глазных яб5
лок), к ядрам Эдингера5Вестфаля и от5
туда через ресничный ганглий к рес5
ничным мышцам (аккомодация) и к
сфинктеру зрачка (сужение зрачка). Во5
локна, идущие к ресничной мышце и
к зрачку, вероятно, не являются одними
и теми же, поскольку зрачковый реф5
лекс на аккомодацию и на свет может
угасать изолированно. Например, при
сифилисе может исчезать реакция зрач5
ка на свет при сохранной реакции на
конвергенцию и аккомодацию (зрачок
До сих пор неизвестно, почему воз5
можна такая ситуация. Кроме того, зра5
чок при этом имеет неправильную фор5
му и сильно сужен, причем сужение
наблюдается очень долго, миоз сохра5
няется и после смерти. По этой причине
было сделано предположение о том, что
нарушение формы зрачка обусловлено
локальным повреждением радужной
оболочки. Недостаточно понятен и синд$
ром Эйди, характеризующийся ненор5
мально медленной, миотонической ре5
акцией зрачка на свет и на темноту, а
также на аккомодацию. (Нарушение пе5
редачи в ресничном
Черепные нервы 109
18
3.23 Центральные пути конвергенции и аккомодации
Межъядерная офтальмоплегия
При унилатеральном повреждении ме5
диального продольного пучка у больно5
го нарушается иннервация ипсилате5
ральной внутренней прямой мышцы
глаза. На рис. 3.24 изображено повреж5
дение левого пучка и паралич левой
внутренней прямой мышцы. Этот па5
ралич возникает несмотря на отсутствие
повреждения ядра или периферического
отрезка нерва, ответственного за эту
мышцу. При этом рефлекторное сокра5
щение мышцы в ответ, например, на
конвергенцию, сохраняется. Когда боль5
ной пытается посмотреть вправо, левый
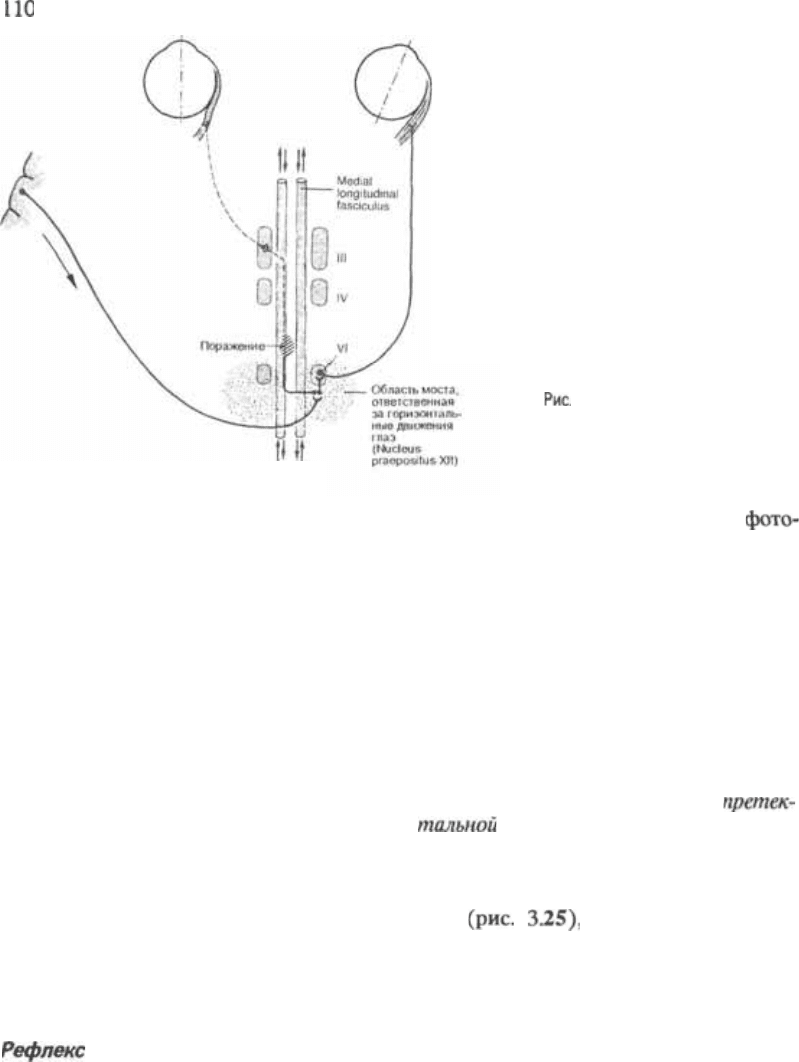
3 Ствол мозга
3.24 Межъядерная офтальмоп*
легия, вызванная поражением меди*
ального продольного пучка.
глаз не движется; в правом глазном
яблоке, иннервируемом отводящим
нервом, возникает моноокулярный нис$
тагм.
Поскольку медиальные продольные
пучки расположены близко друг от дру5
га, один и тот же очаг поражения может
повредить оба эти пучка. При этом ни
одно из глазных яблок не может быть
приведено при горизонтальном отведе5
нии взора. В ведущем глазном яблоке
возникает моноокулярный нистагм. Все
остальные движения глазных яблок воз5
можны; сохраняются и зрачковые ре5
акции.
При унилатеральной межъядерной
офтальмоплегии можно предположить
сосудистую причину ее возникновения,
в особенности у пожилых. Билатераль5
ная межъядерная офтальмоплегия
обычно наблюдается при рассеянном
склерозе.
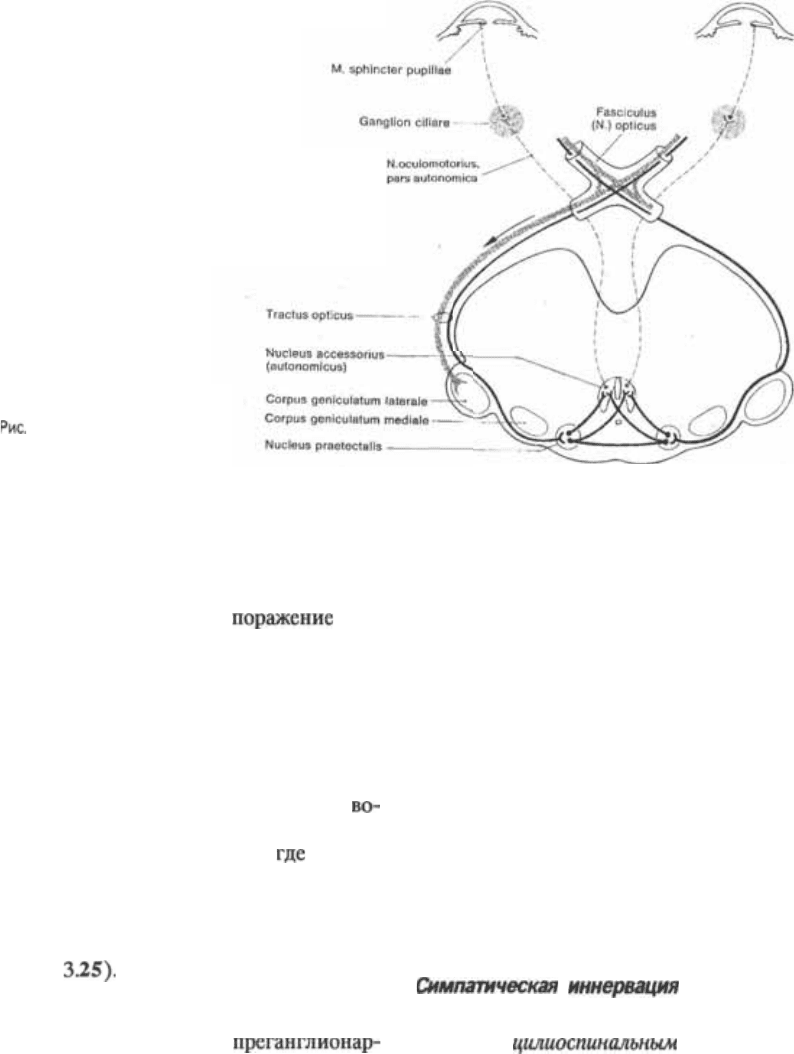
на свет
Когда свет падает на сетчатку, диаметр
зрачка изменяется. Этот зрачковый
рефлекс на свет подобен автоматиче5
ской настройке диафрагмы фотоаппа5
рата: он защищает сетчатку и ее
рецепторы от передозировки световой
энергии и фокусирует изображение
предмета на сетчатку. Этот рефлекс не
затрагивает кору мозга. Поэтому зрач5
ковый рефлекс не осознается.
Афферентные волокна этого рефлек5
са сопровождают зрительный нерв и
зрительный тракт, покидая тракт рядом
с латеральным коленчатым телом в виде
его медиального пучка, направляюще5
гося к верхним бугоркам четверохолмия
и заканчивающегося и ядрах
области. Вставочные нейроны
связывают эти ядра с парасимпатиче5
скими, или автономными добавочными
ядрами Эдингера5Вестфаля обеих сто5
рон обеспечивая синхрон$
ность зрачкового рефлекса на светхвет,
падающий на один глаз, вызывает также
сужение зрачка второго, неосвещенного
глаза. Топография волокон, обеспечи5
вающих зрачковый рефлекс на свет,
служит также объяснением тому, что
разрушение зрительной лучистости или
зрительной коры не влияет на этот реф5
лекс. Нервные волокна, осуществляю5
щие связь с ядрами Эдингера5Вестфаля,
Черепные нервы
111
3.25 Цепь рефлекса на
свет.
еще недостаточно четко выявлены. По5
казано, однако, что разрушение верхних
бугорков не влияет на зрачковый ре5
флекс на свет, тогда как пре5
тектальной области приводит к его уга5
санию, что подтверждает ход афферен5
тных волокон рефлекторной дуги через
эту область. Эфферентные двигательные
волокна происходят из ядер Эдингера5
Вестфаля и вместе с глазодвигательным
нервом входят в глазницу. Здесь пара5
симпатические преганглионарные
локна приобретают самостоятельность
и входят в ресничный узел, сменя5
ются короткими постганглионарными
волокнами. Эти волокна достигают
глазного яблока и иннервируют мышцу,
суживающую зрачок (сфинктер зрачка)
(рис.
Паралич мышцы, суживающей зра5
чок, развивается при повреждении гла5
зодвигательного нерва,
ных волокон или ресничного ганглия.
В результате этого рефлекс на свет уга5
сает, и зрачок расширяется, т. к. сохра5
няется только симпатическая иннерва5
ция. Перерыв афферентных волокон на
протяжении зрительного нерва приво5
дит к исчезновению зрачкового рефлек5
са на свет как на стороне поражения,
так и на противоположной стороне, по5
скольку прерывается сопряженность
этой реакции. Если в этой ситуации,
однако, свет падает на контрлатераль5
ный, непораженный глаз, то рефлекс на
свет возникает с обеих сторон. Ранее
предполагалось, что этот вид мидриаза
является результатом доминирования
симпатических импульсов, вызываю5
щих сокращение мышцы, расширяющей
зрачок (дилататора зрачка). Другое объ5
яснение приведено нами выше.
глаза
Область симпатических ядер, называе5
мая также (реснично5
спинальным) центром, расположена в
боковом роге серого вещества спинного
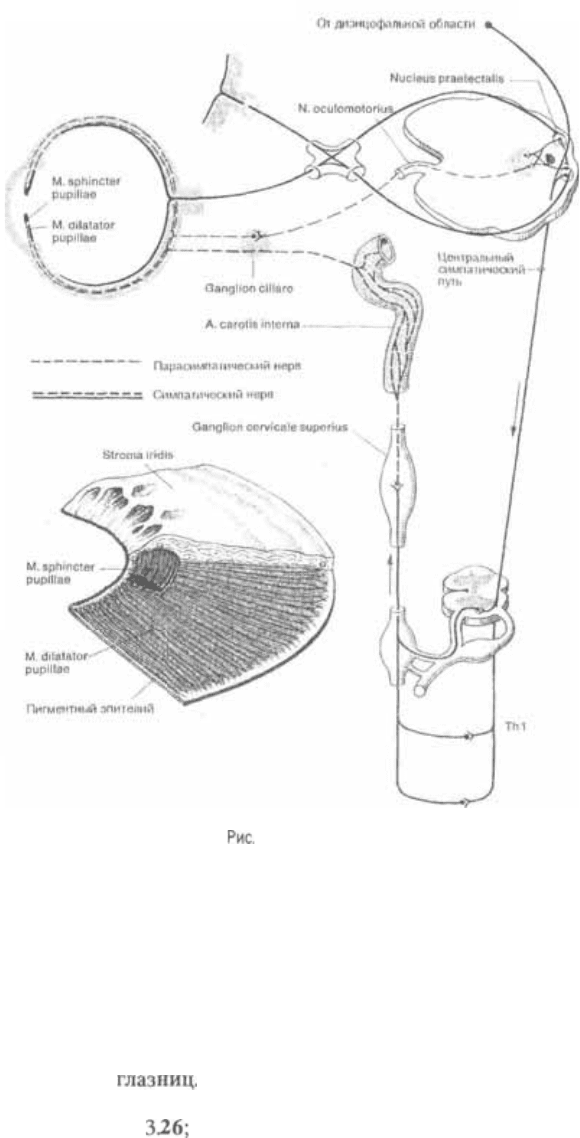
112
3 Ствол мозга
3.26 Парасимпатическая и симпатическая иннервация внутренних мышц глаза.
мозга на уровне от С8 до Т2 сегментов.
Отсюда преганглионарные волокна под5
нимаются к верхнему шейному узлу,
откуда начинаются постганглионарные
волокна. Постганглионарные волокна
присоединяются к внутренней сонной
артерии и поднимаются вместе с ней
до уровня Здесь волокна на5
правляются к мышце, расширяющей
зрачок (рис. см. рис. 3.27). Не
очень ясно, каким образом цилиоспи5
нальный центр получает афферентные
импульсы. Возможно, зрительный тракт
имеет непрямые связи с гипоталаму5
сом. Центральный симпатический путь,
начинающийся там, спускается в сред5
ний мозг, где перекрещивается, и затем
продолжает свой путь через ствол мозга
и шейный отдел спинного мозга к ци5
лиоспинальному центру.
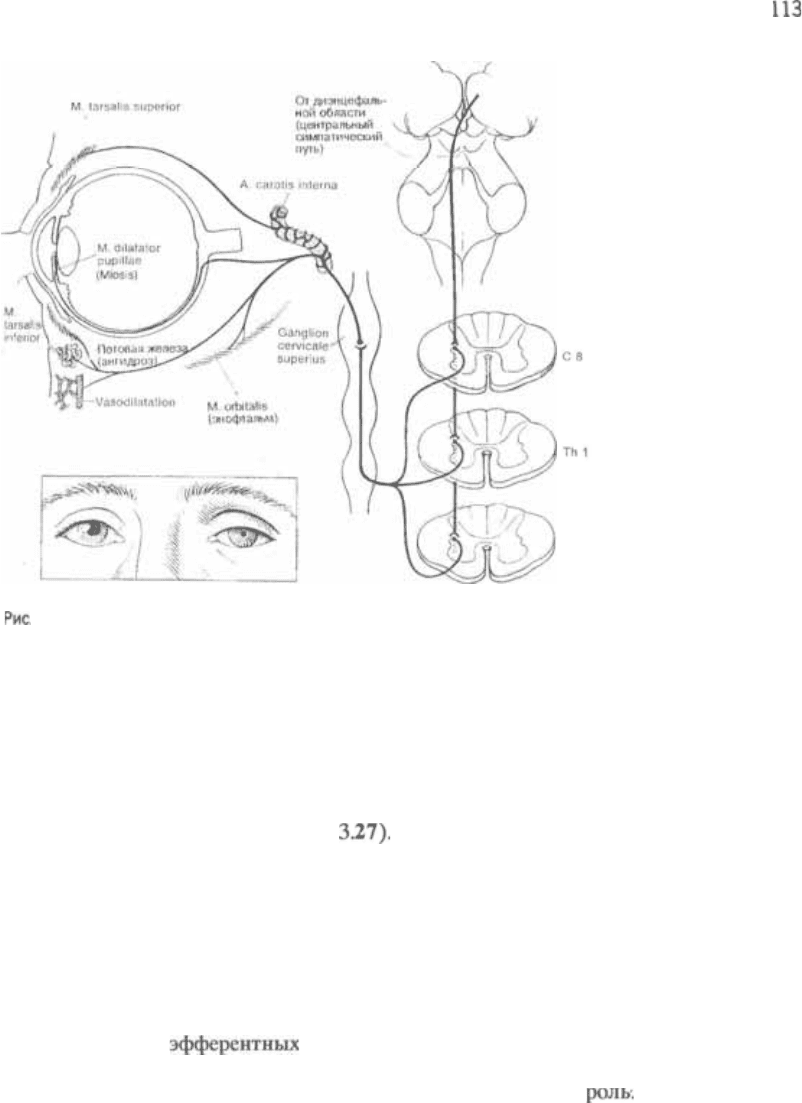
Черепные нервы
Th 2
3.27 Синдром Горнера. Симпатическая иннервация различных структур. Ангидроз и вазодилятация, обусловленные
параличом.
Перерыв импульсов, следующих к
глазу, на уровне центрального симпа5
тического пути, в цилиоспинальном
центре, в верхнем шейном узле или в
области постганглионарных волокон
вызывает синдром Горнера (рис.
Симпатический нерв иннервирует глад5
кие мышечные волокна дилятатора
зрачка, мышцы верхнего и нижнего
хрящей века, а также глазничную мыш5
цу; кроме того, он иннервирует потовые
железы и кровеносные сосуды соответ5
ствующей половины лица. Следователь5
но, при повреждении цилиоспинального
центра или его волокон
на их пути к глазу возникают следую5
щие явления:
1. Сужение глазной щели (паралич тар5
зальных мышц верхнего и нижнего
век);
2. Миоз (паралич дилататора зрачка;
преобладание парасимпатической
иннервации сфинктера зрачка);
3. Энофтальм (паралич глазничной
мышцы);
4. Ангидроз;
5. Вазодилятация соответствующей по5
ловины лица.
Хотя дилататор зрачка и влияет на его
ширину, в последних исследованиях
высказывается предположение о том,
что расширение зрачка при болевых и
эмоциональных реакциях в основном
вызвано торможением парасимпатиче5
ской иннервации, и что симпатическая
иннервация дилататора зрачка играет
незначительную
Как уже указывалось выше, паралич
сфинктера зрачка, вызванный наруше5
нием его парасимпатической иннерва5
114
3 Ствол мозга
ции вследствие поражения ядер Эдин5
гера5Вестфаля, их волокон или реснич5
ного ганглия, сопровождается мидриа5
зом, обусловленным действием полу5
чающего симпатическую иннервацию
зрачка. Разная ширина зрач5
ков глазных яблок называется
Защитный мигательный рефлекс
Внезапное появления предмета прямо
перед глазами немедленно вызывает
рефлекс смыкания век, или рефлекс за$
жмуривания (мигания). Афферентные
волокна этого рефлекса идут от сетчатки
непосредственно к крыше среднего моз5
га и оттуда в составе покрышечно5ядер5
ного пути направляются к ядрам лице5
вых нервов, иннервирующих круговые
мышцы глаз. Если импульсы по
нам пути
нейронов передних рогов шейного от5
дела спинного мозга, то
также поворот головы.
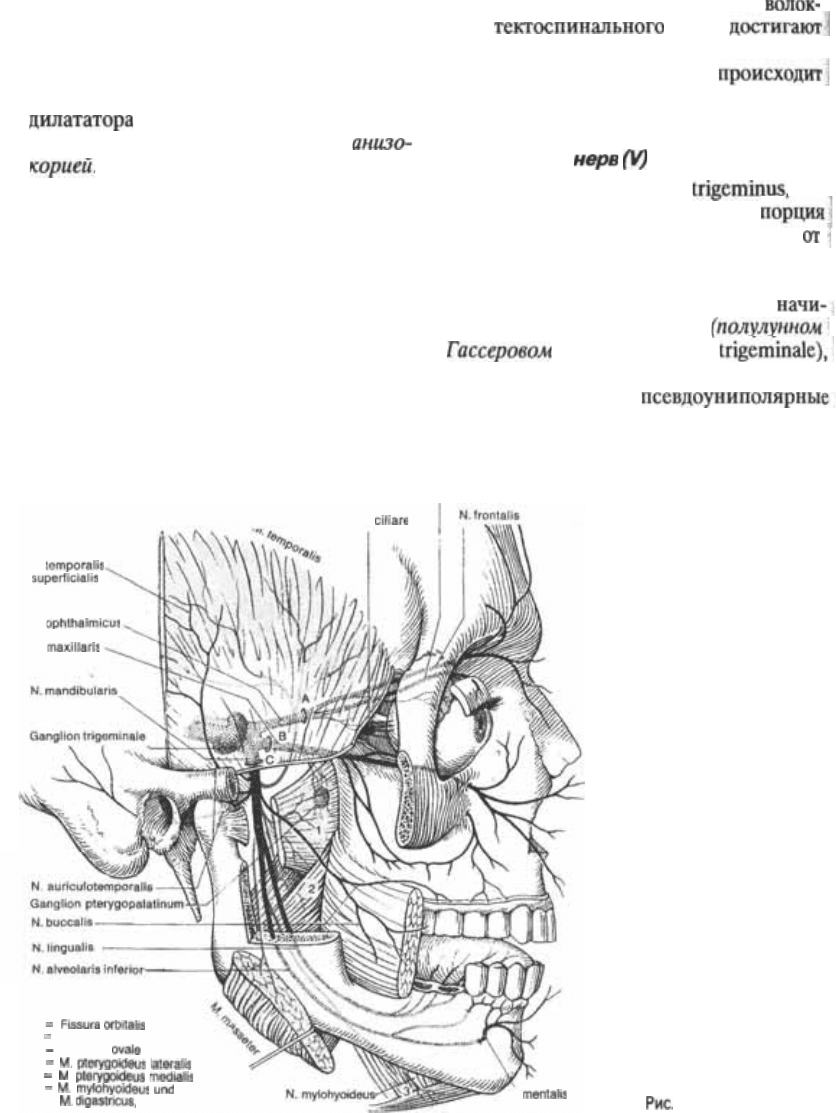
Тройничный
Тройничный нерв, п. яв5
ляется смешанным: его главная
содержит чувствительные волокна
лица, а самая маленькая часть несет
двигательные волокна к жевательным
мышцм. Чувствительная порция
нается в тройничном узле
ганглии, ganglion
который соответствует спинномозгово5
му узлу и содержит
ганглиозные клетки. Периферические
N. nasociliaris
Ganglion
R.
N.
N.
A
В Foramen rofundum
С Foramen
1
2
3
venter anterior
N.
3.28 Тройничный нерв.
Черепные нервы
115
аксоны этих клеток контактируют с ре5
цепторами, воспринимающими при5
косновение, дискриминационную чув5
ствительность, давление, боль и темпе5
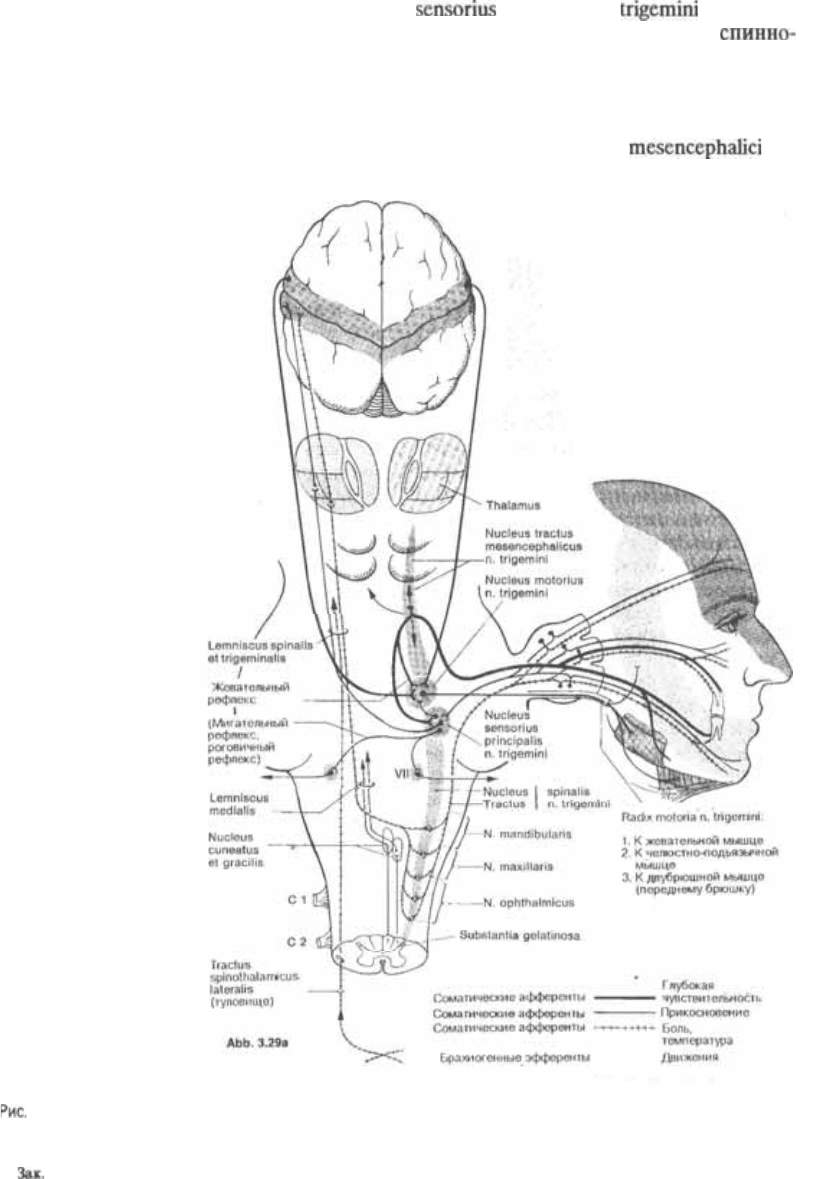
ратуру. Центральные отростки вступают
в варолиев мост, где оканчиваются в
главном чувствительном ядре, nucleus
superior n. (прикос5
новение, дискриминация) и
мозговом ядре, nucleus tractus spinalis n.
trigemini (боль, температура) нерва. Су5
ществует особая точка зрения на ядро
среднемозгового пути тройничного нер$
ва, nucleus tractus n.
3.29а Центральные связи ядер тройничного нерва.
9 136

116
3 Ствол мозга
Вид на двигательную порцию V нерва с
медиальной стороны.
trigemini. Нейроны этого ядра соответ5
ствуют нейронам спинномозгового уз5
ла. Таким образом, это ядро может рас5
сматриваться как ганглий, перемещен5
ный, так сказать, в ствол мозга. Аксоны
его клеток связаны с периферическими
рецепторами мышечных веретен жева5
тельных мышц и с рецепторами, вос5
принимающими давление. Эти три ядра
занимают обширную территорию, изо5
браженную на рис. 329, распространя5
ющуюся от спинного мозга вверх до
среднего мозга.
Гассеров узел расположен в мелкой
выемке (тройничном углублении,
ressio n. trigemini) на вершине пирамид5
ки височной кости снаружи от задне5
бокового отдела кавернозного синуса.
Периферические аксоны нейронов
тройничного узла образуют три основ5
ные ветви:
1. Глазничный нерв, п. ophthalmicus, ко5
торый проходит через верхнюю глаз5
ничную щель.
2. Верхнечелюстной нерв, п.
который проходит через круглое от5
верстие.
3. Нижнечелюстной нерв, п.
который проходит через овальное от5
верстие (см. рис. 3.6).
Периферические ветви этих нервов
показаны на рис. 3.28. Зоны их чувст5
вительной обве5
денные на рис. 3.29 области кожи лба
и лица; слизистую рта и носа,
зубы верхней и нижней челюстей; боль5
шую область твердой мозговой оболоч5
ки передней и средней черепных ямок
Что касается уха, то V нерв снабжает
только переднюю часть наружного уха
и слухового прохода, а также часть ба5
рабанной перепонки. Мочка уха и ос5
тальные части слухового канала получа5
ют чувствительную иннервацию от про5
межуточного нерва (промежуточной
части лицевого нерва), языкоглоточного
и блуждающего нервов. Нижнечелюст5
ной нерв проводит помимо прочих им5
пульсов и проприоцептивные импульсы
от жевательных мышц и неба для конт5
роля за силой прикуса. Область кожной
иннервации тройничного нерва грани5
чит с дерматомами С2 и СЗ спинно5
мозговых нервов. Корешки являются
чисто двигательными и иннервируют
отдельные мышцы шеи, расположенные
между черепом и верхним шейным по5
звонком.
В варолиевом мосту нервные волок5
на, проводящие болевые и температур5
ные импульсы, направляются каудально
в виде спинномозгового тройничного пу$
ти, tractus spinalis n. trigemini, который
оканчивается в спинномозговом ядре
тройничного нерва, распространяю5
щемся вниз до верхнешейного отдела
спинного мозга. Здесь этот путь пред5
ставляет краниальную протяженность
зоны Лиссауэра и желатинозной суб5
станции задних рогов, получающей бо5
левые импульсы от самых верхних шей5
ных сегментов. Каудальная порция
спинномозгового ядра имеет некоторую
соматотопическую организацию. Самая
ее нижняя часть получает болевые во5
локна от глазничного нерва. В более
расположенную часть попа5
дают волокна от верхнечелюстного нер5
ва. Затем следуют волокна от нижнече5
люстного нерва. Волокна от VII (про5

Черепные нервы
117
межуточного нерва) и от IX и X нервов,
проводящие болевые импульсы от уха,
задней трети языка, глотки и гортани,
присоединяются к спинномозговому
пути тройничного нерва (см. рис. 3.41
и 3.42). Средний сегмент (интерполи5
рованная часть) и краниальный сегмент
(ростральная часть) спинномозгового
ядра, возможно, получают афферентные
волокна, проводящие импульсы давле5
ния и прикосновения. Считается, что
средний сегмент получает болевые во5
локна, начинающиеся в пульпе зубов.
Необходимо, однако, дальнейшее уточ5
нение функции данной части ядра.
Аксоны вторых нейронов спинно5
мозгового ядра расходятся веером при
переходе на противоположную сторону,
где они проходят через покрышку ва5
ролиева моста и поднимаются к тала5
мусу вместе с боковым
ным путем. Эти волокна оканчиваются
в вентральном задне5срединном ядре
зрительного бугра (см. рис. 3.29).
Основное чувствительное ядро трой$
ничного нерва занимает ограниченную
область дорсолатеральной части по5
крышки моста. Оно получает афферен5
тные импульсы прикосновения, диск5
риминации и давления, которые в спин5
ном мозгу проводятся задними кана5
тиками. Аксоны вторых нейронов этого
ядра также переходят на противополож5
ную сторону и идут вместе с медиаль5
ной петлей до вентрального заднемеди5
ального ядра таламуса. Третьи нейроны
тройничных путей, расположенные в та5
ламусе, посылают свои аксоны через
заднюю ножку внутренней капсулы до
нижней трети постцентральной извили5
ны (см. рис. 329а и 1.20).
Двигательная, или
тройничного нерва, radix
имеет свое ядро в покрышке
моста, nucleus n. trigemini. Оно
расположено медиальнее основного чув5
ствительного ядра. Двигательная пор5
ция выходит из черепа вместе
с нижнечелюстным нервом и иннерви5
жевательную, височную, латераль5
ную и медиальную крыловидные, че5
люстно5подъязычную мышцы, перед5
нее брюшко двубрюшной и
мышцу, натягивающую мягкое небо
(рис. Двигательное ядро получа5
ет центральные стимулы по корково5
ядерному пути. Этот путь
начинается от больших пирамидных
клеток нижней трети прецентральной
извилины (см. рис. 3.29а, 22 820).
Импульсы поступают преимущественно
с противоположной стороны. Но по5
скольку часть их приходит с ипсилате5
ральной стороны, одностороннее пре5
рывание надьядерных тройничных пу5
тей не вызывает заметной слабости же5
вательных мышц. В противоположность
этому, поражение ядра или перифери5
ческой части двигательной порции нер5
ва вызывает развитие вялого паралича
соответствующих жевательных мышц,
за которым следует их атрофия. Такой
односторонний паралич легко может
быть распознан по отсутствию сокра5
щений жевательной мышцы при по5
пытке сжать челюсти. Если попросить
больного открыть рот и выдвинуть под5
бородок вперед, то заметно, что подбо5
родок отклоняется в сторону паралича
из5за преобладания действия крыловид5
ных мышц другой стороны. На пара5
лизованной стороне также отсутствует
рефлекс с жевательной мышцы (жева$
тельный рефлекс). В норме жевательная
мышца сокращается в ответ на удар по
ней неврологическим молоточком при
открытом рте.
Сенсорные импульсы от слизистой
оболочки глаза проводятся по глазнич5
ному нерву до основного чувствитель5
ного ядра. Здесь они переключаются на
другие нейроны, представляющие аф5
ферентную часть дуги реф$
лекса и связанные с ядром лицевого
нерва на этой же стороне. Эфферентная
часть рефлекторной дуги представлена
9*