Чупахина Г.Н. Система аскорбиновой кислоты растений
Подождите немного. Документ загружается.

КАЛИНИНГРАДСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Г.Н. Чупахина
СИСТЕМА АСКОРБИНОВОЙ
КИСЛОТЫ РАСТЕНИЙ
Калининград
1997
КАЛИНИНГРАДСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Г.Н. Чупахина
СИСТЕМА АСКОРБИНОВОЙ
КИСЛОТЫ РАСТЕНИЙ
Монография
Калининград
1997
Чупахина Г.Н. Система аскорбиновой кислоты растений: Монография. -
Калинингр. ун-т. - Калининград, 1997. - 120 с. - ISBN 5-88874-063-2.
В монографии дана характеристика системы аскорбиновой кислоты растений,
включающей аскорбиновую, дегидроаскорбиновую и дикетогулоновую кислоты, а
также ферменты, окисляющие и восстанавливающие аскорбат. Приведены результаты
исследования действия полихроматического и монохроматического света на
компоненты системы, обсуждается вопрос о фоторецепторе светозависимого
синтеза
аскорбиновой кислоты. Рассматриваются пути биосинтеза аскорбата, компартментация
данного процесса и субклеточная локализация кислот системы аскорбиновой кислоты.
Исследована зависимость биосинтеза указанных кислот от фотосинтеза и дыхания.
Впервые исследовалась дикетогулоновая кислота как компонент системы аскорбиновой
кислоты. Приводится метод получения альбиносных проростков, их физиолого-
биохимическая характеристика, рекомендация использования в качестве модели для
изучения
светозависимых процессов не связанных с фотосинтезом.
Для физиологов, биохимиков, преподавателей вузов, аспирантов и студентов.
Иллюстр. 30. Библиогр.: 461 назв.
Рецензент - зав. отделом химии фотосинтеза Института биоорганической химии и
нефтехимии АН Украины, чл.-кор. АН Украины А.А. Ясников.
Печатается по решению редакционно-издательского Совета Калининградского
государственного университета.
ISBN 5-88874-063-2 ©
Калининградский государственный
университет, 1997
Галина Николаевна Чупахина
СИСТЕМА АСКОРБИНОВОЙ КИСЛОТЫ РАСТЕНИЙ
Монография
Лицензия № 020345 от 27.12.1991 г.
Редактор Л.Г. Ванцева.
Оригинал-макет подготовлен Д.В. Голубиным.
Подписано в печать 25.12.1996 г. Формат 60×90 1/16.
Бумага для множительных аппаратов. Усл. печ. л. 7,5.
Уч.-изд. л. 8,0. Тираж 200 экз. Заказ 92.
Калининградский
государственный университет,
236041, г. Калининград, ул. А. Невского, 14.
3
ВВЕДЕНИЕ
Аскорбиновая кислота - уникальное полифункциональное соединение.
Обладая способностью обратимо окисляться и восстанавливаться, она
принимает участие в важнейших энергетических процессах растительной клетки
- фотосинтезе [341, 378] и дыхании [451, 362]; является признанным
антиоксидантом [8]. Несомненно ее участие в процессах роста, цветения,
вегетативной и репродуктивной дифференциации [230], в водном обмене [151],
регуляции ферментативной активности [46], стимуляции реакций метаболизма,
связанных с обменом нуклеиновых
кислот и синтезом белка [207, 365], в
защитных реакциях растений [200, 2].
Особый интерес для биотехнологии представляет факт накопления
аскорбиновой кислоты в культуре растительных тканей и активации их роста
аскорбатом [288, 295]. В связи с этим знание условий, формирующих пул
аскорбиновой кислоты зеленых растений, необходимо не только для
программированного получения высоковитаминных растительных продуктов,
что само по себе
является важным для решения проблемы качества пищи, но и
для понимания механизмов, определяющих продуктивность растений.
Возросший интерес к аскорбиновой кислоте как к лекарственному препарату
[257] обусловлен противовирусным [395] и противоопухолевым действием
аскорбиновой кислоты и ее дериватов [450], а также авторитетной рекламой
дважды лауреата Нобелевской премии Л.Полинга [116]. При определении
витаминной ценности растительного сырья
витамин С определяется как сумма
взаимопревращающихся восстановленной и окисленной форм аскорбиновой
кислоты [152]. Однако дегидроаскорбиновая кислота не всегда переходит в
аскорбиновую, так как разрыв ее лактонового кольца ведет к необратимому
образованию дикетогулоновой кислоты, не обладающей витаминной актив-
ностью и практически не исследованной у растений. Поэтому для всесторонней
оценки роли аскорбиновой кислоты
в метаболизме растений необходимо
одновременное исследование всей системы аскорбата, включающей
вышеназванные органические кислоты, а также ферментные системы,
способствующие окислению и восстановлению аскорбиновой кислоты. Именно
4
такие подходы к исследованию витамина С [258, 430] являются наиболее
перспективными.
Со времени открытия аскорбиновой кислоты интерес к этому соединению не
проходит, хотя в последние годы он значительно снизился. Но это не говорит о
том, что решены все проблемы, связанные с аскорбатом. Они есть. В частности,
до сих пор не выяснены пути синтеза
аскорбиновой кислоты, не до конца ясна ее
роль в жизни автотрофных организмов, являющихся продуцентами и
поставщиками витамина С для человека. Решение этих вопросов требует нового
уровня исследований, чему, несомненно, может служить изучение аскорбиновой
кислоты как компонента системы органических кислот: аскорбиновая ↔
дегидроаскорбиновая → дикетогулоновая кислота.
В предлагаемой работе обобщены литературные
данные и результаты
собственных исследований автора, касающиеся путей биосинтеза аскорбиновой
кислоты и субклеточной локализации аскорбиновой, дегидроаскорбиновой и
дикетогулоновой кислот. Рассматривается действие полихроматического и
монохроматического света на компоненты системы, зависимость биосинтеза
анализируемых кислот от фотосинтеза и дыхания. Для решения этих вопросов
использовались данные по биосинтезу кислот в проростках ячменя с
измененным пигментным
составом, в экспериментально полученных
альбиносных проростках, а также при ингибировании и стимуляции дыхания
интермедиатами цикла Кребса.
5
СПИСОК СОКРАЩЕНИЙ
АК - аскорбиновая кислота
АО - аскорбатоксидаза
ГН - глютатион восстановленный
ГS-SГ - глютатион окисленный
ДАК - дегидроаскорбиновая кислота
ДКГК - дикетогулоновая кислота
ДКС - дальний красный свет
2,4 - ДНФ - 2,4 - динитрофенол
ИУК - индолилуксусная кислота
КС - красный свет
МДАК - монодегидроаскорбиновая кислота
НАД - никотинамидадениндинуклеотид
НАДФН+Н
+
- никотинамидадениндинуклеотидфосфат восстановленный
ОПФЦ - окислительный пентозофосфатный цикл
ПААГ - полиакриламидный гель
СМ - стрептомицин
УФ - ультрафиолет
ФАР - физиологически активная радиация
Ф
дк
- фитохром с максимумом поглощения в области дальнего красного
света около 730 нм
ФС I - фотосистема I
ФС II - фотосистема II
Хл - хлорофилл
ЦТК - цикл трикарбоновых кислот
ЭТЦ - электрон-транспортная цепь
6
Глава 1. КИСЛОТЫ И ФЕРМЕНТЫ СИСТЕМЫ
АСКОРБИНОВОЙ КИСЛОТЫ
В 1927 году A.Сцент-Гиорги впервые выделил из некоторых растений и
коры надпочечников неустойчивое вещество с сильно выраженными
восстановительными свойствами, которое сначала назвал гексуроновой
кислотой, затем - аскорбиновой, учитывая ее антискорбутное (скорбут - цинга)
действие. Химическая формула АК была предложена в 1933 году Хирстом и
независимо от
него Эйлером, а в конце 1933 г. ее правильность окончательно
доказана Михеелем. Химический синтез АК осуществлен Рейхштейном в 1932
г., когда структура ее еще не была известна. В 1933 г. Рейхштейн и Хаворс с
Хирстом опубликовали методы синтетического получения АК. История
открытия и изучения АК подробно описана Б.А. Кудряшовым [69].
В химическом отношении
АК является лактоном 2,3-диэнол-l-гулоновой
кислоты с эмпирической формулой С
6
Н
8
О
6
. Она обладает сильно выраженными
восстанавливающими свойствами благодаря наличию в молекуле диэнольной
группы:
С ОН
С ОН
Окисляясь, АК легко переходит в дегидроформу:
O O
C C
НО-С
О = С
НО-С О - 2H О = С О
H-C + 2H
H-C
HO-C-H HO-C-H
СH
2
-OH СH
2
-OH
L-аскорбиновая Дегидроаскорбиновая
кислота кислота
(восстановленная форма) (окисленная форма)
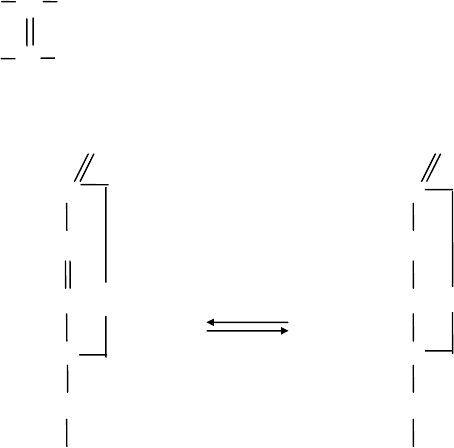
Этот процесс обратимый, и ДАК может вновь восстановиться до l-AK.
Окисление АК до ДАК происходит в результате отдачи двух протонов и
двух электронов. При окислении АК в особых условиях (УФ свет, температура
жидкого воздуха) за счет отдачи только одного протона возникает свободный
радикал - монодегидроаскорбиновая кислота. Это нестабильная,
реакционноспособная форма АК.
7
O O O O
C
C C C
НО-С
HO-С O-С O=С
НО-С
О -H
+
О-С О -H
+
О-С О -2e О=С О
H-C H-C H-C H-C
HO-C-H HO-C-H HO-C-H HO-C-H
СH
2
-OH СH
2
-OH СH
2
-OH СH
2
-OH
L-аскорбиновая Монодегидро- Анион дегидро- Дегидроаскор-
новая кислота аскорбиновая аскорбиновой биновая кис-
кислота кислоты лота
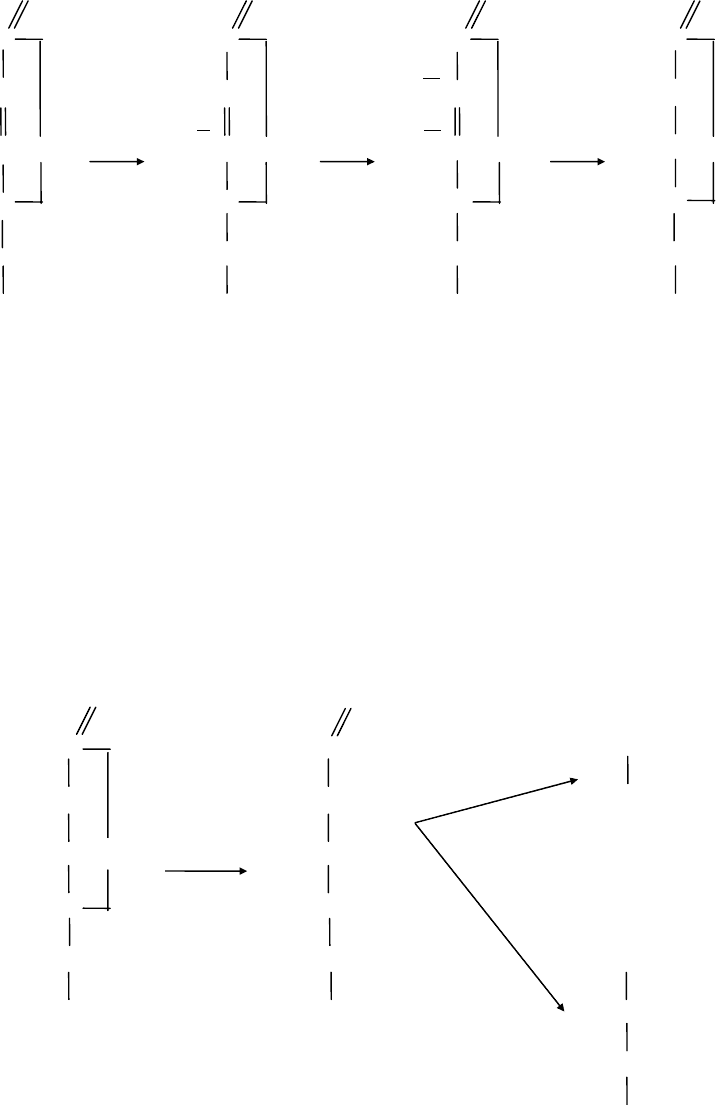
Переход l-АК в ДАК двухэтапный. На первом этапе две молекулы l-AK дают
начало двум молекулам монодегидроаскорбиновой кислоты. Далее они
дисмутируют: одна молекула превращается в ДАК, другая - в l-АК [60].
Лактоновое кольцо в молекуле l-AK стабильно, а в ДАК легко гидролизует с
образованием кислоты с открытой цепью - 2,3-дикетогулоновой кислоты [84].
Последняя может
подвергаться окислительному распаду под действием
гипоиодида натрия с образованием щавелевой и треоновой кислот:
O O
C C - OH COOH
О = С С = O COOH
О = С О +H
2
O С = О Щавелевая
кислота
H - C H - C - OH
HO
- C - H HO - C - H COOH
СH
2
- OH СH
2
- OH H - C - OH
HO - C - H
CH
2
OH
Дегидроас- 2,3-дикето-
корбиновая l-гулоновая L-треоновая
кислота кислота кислота
8
Введенная экзогенно дикетогулоновая кислота при освещении хлоропластов
фотоокисляется [453]. Под влиянием окисляющих АК ферментов происходит
декарбоксилирование ДАК и ДКГК с образованием l-ксилоновой и l-ликсоновой
кислот.
У растений, аккумулирующих щавелевую кислоту (шпинат, бегония,
кислица), окисление введенной 1-
14
С-АК или 1-
14
С-ДАК происходит с
образованием щавелевой и винной кислот. В проростках ячменя, которые не
накапливают щавелевую кислоту, включение метки в последнюю происходит
слабо. Отмечено, что метка из 1-
14
C-ДАК легко включалась в щавелевую
кислоту шпината и кислицы, а 1-
14
С-ДКГК очень медленно метаболизировалась
в щавелевую кислоту. Вероятно, для образования щавелевой кислоты
существенно наличие лактонового кольца в предшественнике [453].
У герани также два первые атома углерода АК включаются в молекулу
щавелевой кислоты, а С
4
-фрагмент - в винную кислоту [445, 448]. Однако судьба
двух первых атомов углерода АК может быть иной. Например, у винограда
отрезок цепи с первого по четвертый атом углерода включается в l-винную
кислоту, а оставшийся С
2
-фрагмент повторно вовлекается в цикл гексоз и
продуктов их метаболизма. [445, 383]. При этом углерод первого атома АК
может высвобождаться в форме CO
2
[272].
Изолированные листья Parthenocissus quinquefolia L метаболизируют АК с
образованием l-треоновой и винной кислот. Показано, что треоновая кислота не
является промежуточным метаболитом обмена аскорбата, превращающегося в
винную кислоту [271]. Такими метаболитами могут быть l-идониновая, 2-кето-l-
идониновая и 5-кето-l-идониновая кислоты. В молодых тканях винограда вве-
денная АК превращается в 5-кето-
l-идониновую кислоту через 2-кето-l-
идониновую и l-идониновую кислоты. Из С
4
-фрагмента 5-кето-l-идониновой
кислоты далее синтезируется l-(+)винная кислота. Считается [318], что именно l-
АК. является физиологическим интермедиатом на пути биосинтеза винной
кислоты в листьях винограда. Радиоактивность от 5-кето-l(6-
14
С)идониновой
кислоты обнаруживалась в сахарах и нерастворимом осадке [381, 382].
Следовательно, обмен АК в растениях связан с органическими кислотами и
углеводами, которые в одних условиях являются субстратом в биосинтезе АК, а
в других для их образования используются фрагменты молекулы АК,
полученные после ее деградации.
Таким образом, АК в растительных тканях может присутствовать
в
восстановленной форме, окисленной и в виде нестабильной
монодегидроаскорбиновой кислоты. Восстановленная и окисленная АК
находятся в свободном состоянии. Известны три связанные формы АК:
аскорбиген - соединение АК с полипептидом; комплекс АК с витамином Р и,
вероятно, еще с каким-то неизвестным веществом; третья связанная форма АК -
это соединение АК с нуклеиновой
кислотой через посредство минерального
железа [158]. Обычно в растениях преобладает восстановленная форма АК. Накоплению
ДАК препятствует ее нестабильность (особенно при рН выше 5), поэтому при
измельчении тканей наблюдается быстрая потеря ДАК. Тем не менее измеримые
количества ДАК присутствуют во многих тканях. Содержание окисленной
формы АК в разных условиях неодинаково и иногда достигает высоких