Balian E.V., L?v?que C., Segers H., Martens K. (Eds.) Freshwater Animal Diversity Assessment
Подождите немного. Документ загружается.

FRESHWATER ANIMAL DIVERSITY ASSESSMENT
Global diversity of cumaceans & tanaidaceans (Crustacea:
Cumacea & Tanaidacea) in freshwater
D. Jaume Æ G. A. Boxshall
Springer Science+Business Media B.V. 2007
Abstract Cumacea and Tanaidacea are marginal
groups in continental waters. Although many eury-
haline species from both groups are found in estuaries
and coastal lagoons, most occur only temporarily in
non-marine habitats, appearing unable to form stable
populations there. A total of 21 genuinely non-mar ine
cumaceans are known, mostly concentrated in the
Ponto-Caspian region, and only four tanaids have
been reported from non-marine environments. Most
non-marine cumaceans (19 species) belong in the
Pseudocumatidae and appear restricted to the Caspian
Sea (with salinity up to 13 %) and its peripheral
fluvial basins, including the northern, lower salinity
zones of the Black Sea (Sea of Azov). There are nine
Ponto-Caspian genera, all endemic to the region.
Only two other taxa (in the family Nannastaci dae)
occur in areas free of any marine–water influence, in
river basins in North and South America. Both seem
able to survive in waters of raised salinity of the
lower reaches of these fluvial systems; but neither has
been recorded in full salinity marine environments.
The only non-marine tanaidacean thus far known
lives in a slightly brackish inland spring in Northern
Australia. The genus includes a second species, from
a brackish-water lake at the Bismar ck Archipelago,
tentatively included here as non-marine also. Two
additional species of tanaidaceans have been reported
from non-marine habitats but both also occur in the
sea.
Keywords Freshwater Global assessment
Species richness Peracarida Crustacea
Introduction
Comprising about 1,300 and 900 marine species,
respectively, the Cumacea and Tanaidacea are only
marginal groups in continental waters. Altho ugh
many euryhaline species in both taxa are found in
estuaries and coastal lagoons, most occur only
temporarily in non-marine habitats, appearing unable
to form stable populations there. Only 21 genuinely
non-marine cumaceans are known, most of which
occur in the Ponto-Caspian region, whereas just four
tanaids have been reported in non-marine habitats.
Both groups are orders of peracarid crustaceans that
are mainly adapted to a fossorial life-style in non-
consolidated marine sediments, especially in deep
Guest editors: E. V. Balian, C. Le
´
ve
ˆ
que, H. Segers &
K. Martens
Freshwater Animal Diversity Assessment
D. Jaume (&)
IMEDEA (CSIC-UIB), Instituto Mediterra
´
neo de Estudios
Avanzados, C/ Miquel Marque
`
s 21, Esporles, Illes Balears
07190, Spain
e-mail: d.jaume@uib.es
G. A. Boxshall
Department of Zoology, The Natural History Museum,
Cromwell Road, London SW7 5BD, UK
e-mail: g.boxshall@nhm.ac.uk
123
Hydrobiologia (2008) 595:225–230
DOI 10.1007/s10750-007-9018-0

waters, although they can appear regularly in night-
time plankton hauls in shallow waters.
The characteristic body form of a cumacean
consists of a large, variably inflated cephalothorax
incorporating the first 3 (of 8) thoracic somites, plus
an elongate, narrow abdomen terminating in a pair of
long and slender uropod s. The cephalothorax displays
a pair of frontal extensions, the pseudorostral lobes,
which converge medially in most instances, whereas
its lateral portions act as paired branchial chambers
accommodating the respiratory epipodites of the first
maxillipeds (see below). All thoracopods except the
first, second and eighth are primitively biramous. The
first pair (=first maxillipeds) is characteristic, pos-
sessing a respiratory coxal epipodite provided with
digitiform extensions in addition to a narrow frontal
extension, which together with the corresponding
cephalothoracic pseudorostral lobe, forms a branchial
siphon (exhalant canal) for the corresponding bran-
chial chamber. The abdomen comprises six free
pleomeres and a free telson, although in some
families the latter is incorporated into the sixth
pleomere forming a pleotelson. Apart from the
uropods on the last pleomere, there are up to five
pairs of pleopods in males, but a maximum of only
one pair in females. Reduction in number of pairs of
pleopods is common. All these limbs are originally
biramous, with a 2-segmented exopod and a uniseg-
mented endopod; the endopod of the uropod can be
up to 3-articulate. Cumaceans are primarily deposit
feeders, although some are apparently predators of
foraminifers and other crustaceans. Most live half-
buried in soft sediments.
General morphological characteristics for the
order Tanaidacea include: a small cephalothorax
incorporating the first two thoracic somites, six free
thoracic somites, five abdominal somites bearing
pleopods, and a pleotelson with a pair of uropods. All
thoracopods except the third (=first pereiopod) of
most apseudomorphs, and some other pereiopods of
the manca stages of the genus Kalliapseudes, are
uniramous. The maxillipeds (=first thoracopods)
possess a respiratory coxal epipodite, which is
concealed under the lateral margin of the cephalo-
thorax (branchial cavity). The second pair of
thoracopods is prehensile, displaying a chelate distal
portion (‘‘chelipeds’’). The pleopods and uropods are
basically biramous with 2-segmented exopods and
unisegmented endopods, although both rami of the
uropods can be multi-articulate, due to the display of
cuticular annulations. Tanaidaceans are primarily
tube or tunnel dwellers, and are generally considered
to be deposit feeders.
Species diversity, distribution and historical
processes
Non-marine cumaceans belong to two of the eight
recognised families: Pseudocumatidae Sars and Nan-
nastacidae Bate. Most non-marine species (19) are
pseudocumatids and their distribution is focused
around the Caspian Sea (maximum salinity 13%)
and its peripheral fluvial basins, including the north-
ern, lower salinity zones of the Black Sea (Sea of
Azov) (see Tables 1 and 3; Fig. 1). They represent
nine genera, all endemic to the region, although the
taxonomic status of some genera is equivocal (e.g.
Charsarocuma; see comments by Sars (1914: 32) on
its presumed synonymy with Schizoramphus ) and
their validity should be tested. The natural distribu-
tion of these taxa within the Ponto-Caspian region is
difficult to ascertain since dispersal via artificial
canals and reservoirs, by shipping, or even by
deliberate introduction as fish food, may have had a
profound effect (see Ba
˘
cescu & Petrescu, 1999, and
references therein). Stenocuma graciloides has
recently been reported from the Gulf o f Finland
(Baltic Sea), where it may have been transported by
ships passing through the Volga-Baltic waterway
from its North Caspian home (Antsulevich, 2005).
The presumed deliberate introduction of Stenocuma
gracilis, Pterocuma pectinata and Schizorhamphus
scabriusculus into the Aral sea, to serve as fish food
(Karpevitch, 1960; quoted in Ba
˘
cescu & Petrescu,
1999), seems not to have succe eded (Nikolay Aladin,
pers. comm.). Apart from these Caspian pseudocu-
matids, only two other taxa (from the Nannastacidae)
occur in areas free of any marine-water influence, in
river basins in North and South America. Both seem
able to survive in waters of raised salinity of the
lower reaches of these fluvial systems (see Tables 1
and 3; Fig. 1), but neither has been recorded in full
salinity marine environments. These two monotypic
genera are endemic to their respective river basins.
Sars (1914) considered that the Caspian Cumacea
were derived from a single ancestral form originating
from the Mediterranean, probably belonging to the
226 Hydrobiologia (2008) 595:225–230
123

marine genus Pseudocuma Sars, 1865, which
includes three Mediterranean species, one of which
is also present in the Black Sea. The Caspian genus
Stenocuma is considered to be a subgenus of
Pseudocuma by some authors (Ba
˘
cescu, 1992).
Dumont (2000: 186) believed that Caspian cuma-
ceans were derived from ancestral forms that lived in
estuaries and tidal zones of rivers that discharged into
Table 1 Global diversity of non-marine Cumacea (distribution of Ponto-Caspian taxa after Ba
˘
cescu (1992) and Antsulevich (2005))
Order Cumacea Distribution
Family Pseudocumatidae G. O. Sars, 1878
Genus Carinocuma Mordukhai-
Boltovskoi & Romanova, 1973
C. birsteini Mordukhai-Boltovskoi &
Romanova, 1973
Caspian Sea
Genus Caspiocuma G. O. Sars, 1900
C. campylaspoides (G. O. Sars, 1897) Caspian Sea; Volga, Don, Bug and Dniestr river basins
Genus Charsarocuma Derzhavin, 1912
C. knipowitschi Derzhavin, 1912 Caspian Sea; pre-delta region of Volga
Genus Hyrcanocuma Derzhavin, 1912
H. sarsi Derzhavin, 1912 Caspian Sea
Genus Stenocuma G. O. Sars, 1900
S. gracilis (G. O. Sars, 1894) Caspian Sea; Volga
S. graciloides (G. O. Sars, 1894) Caspian Sea; Estuaries of Volga, Don, Dniestr and Danube; Black Sea (Azov);
Gulf of Finland (Baltic Sea)
S. tenuicauda (G. O. Sars, 1894) Caspian Sea; Volga
S. diastyloides (G. O. Sars, 1897) Caspian Sea
S. cercarioides G. O. Sars, 1894 Caspian Sea; Volga, Don, Bug and Dniestr river basins; Black Sea
S. laevis (G. O. Sars, 1914) Caspian Sea
Genus Pterocuma G. O. Sars, 1900
P. pectinata (Sowinski, 1893) Caspian Sea; Volga, Danube and Dniestr river basins; Black Sea
P. rostratum (G. O. Sars, 1894) Caspian Sea; Volga; estuaries of Dniepr, Bug and Danube; Black Sea
P. sowinskyi (G. O. Sars, 1894) Caspian; Volga; delta of Don; Black Sea
P. grandis G. O. Sars, 1914 Caspian Sea
Genus Schizorhamphus Ba
˘
cescu, 1992
S. bilamellatus (G. O. Sars, 1894) Caspian Sea; Volga
S. eudorelloides (G. O. Sars, 1894) Caspian Sea (up to 264 m depth); river mouths of Danube, Dniestr and Prut;
Black Sea
S. scabriusculus (G. O. Sars, 1894) Caspian Sea; Danube, Dniestr, Bug and Dniepr rivers
Genus Strauchia Czerniavsky, 1868
S. taurica Czerniavsky, 1868 Caspian Sea
Genus Volgocuma Derzhavin, 1912
V. telmatophora Derzhavin, 1912 Caspian Sea; Volga; Black Sea
Family Nannastacidae Bate, 1866
Genus Almyracuma Jones & Burbanck,
1959
A. proximoculi Jones & Burbanck, 1959 Intertidal freshwater springs at Cape Cod, and limnetic zone of Lower Hudson river
(latter 1–30%; Simpson et al., 1985), NE U.S.A.
Genus Claudicuma Roccatagliata, 1981
C. platense Roccatagliata, 1981 Rı
´
o de la Plata (Argentina), from Buenos Aires (0.5%) to Punta del Indio (1.8–7.0%)
(Roccatagliata, 1991)
Hydrobiologia (2008) 595:225–230 227
123
the (proto-) Mediterranean before the closure of the
Sarmatian Basin, a vanished Miocene brackish lake
that covered the entire Ponto-Caspian region from
14.5 to 8.3 Myr ago. Their osmoregulatory abilities
would have preadapted them to life in the brackish
Sarmatian lake.
The only truely non-marine tanaidacean known is
Pseudohalmyrapseude s aquadulcis (Parapseudidae)
which lives in a slightly brackish inland spring in
Northern Australia (see Tables 2 and 3; Fig. 1; Larsen
& Hansknecht, 2004). The genus includes a second
species, P. mussauensis, from a brackish-water lake
in the Bismarck Archipelago; this species is tenta-
tively included here as non-marine, since the genus
has not been recorded yet in fully marine environ-
ments, and Shiino (1965) was rather vague in his
description of the salinity regime of the lake where the
species was discovered (see Tables 2 and 3; Fig. 1).
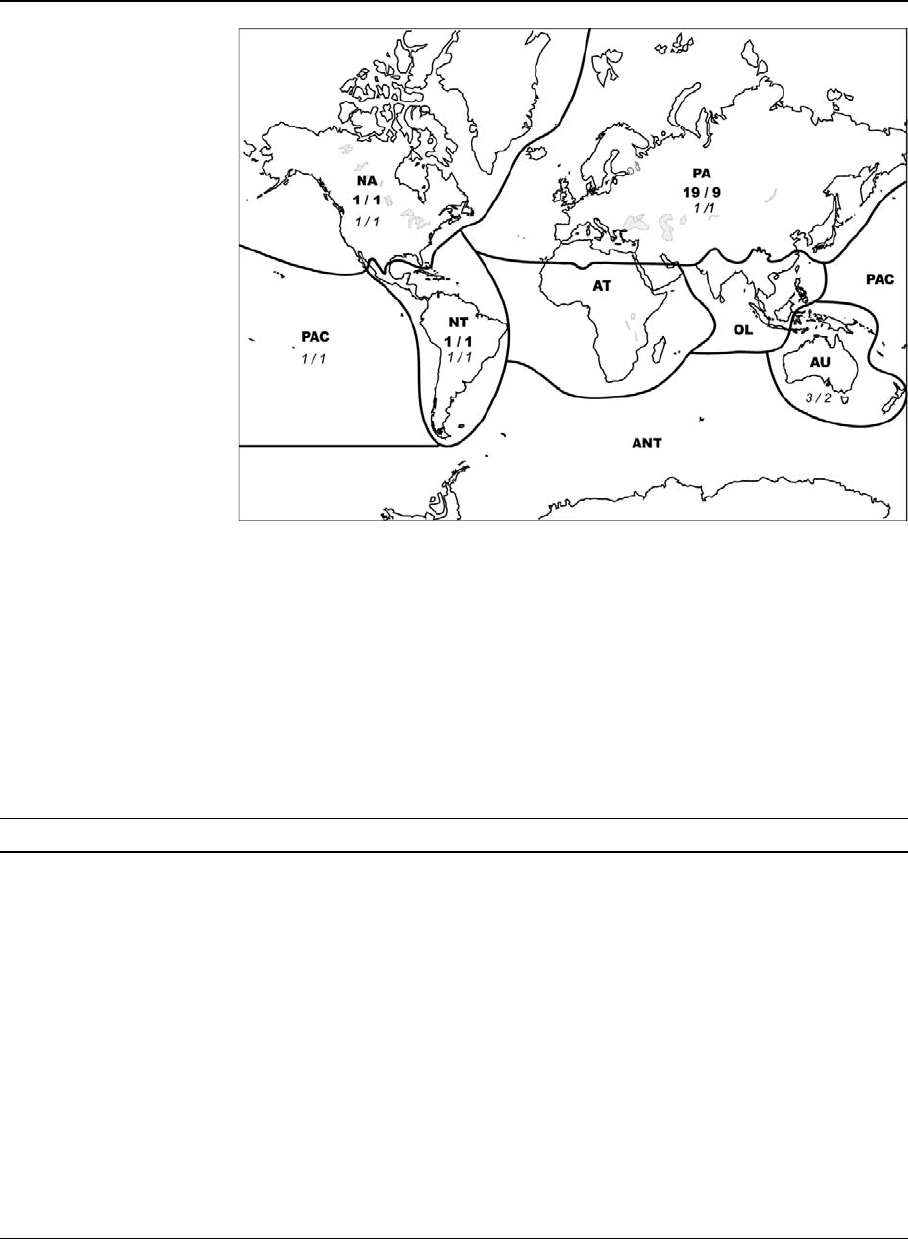
Fig. 1 Total species and
genus numbers of Cumacea
(Bold) and Tanaidacea
(italics) per zoogeographic
regions (Species number/
Genus number). PA:
Palaearctic, NA: Nearctic,
NT: Neotropical, AT:
Afrotropical, OL: Oriental,
AU: Australasian, PAC:
Pacific Oceanic Islands,
ANT: Antarctic
Table 2 Global diversity of non-marine Tanaidacea
Order Tanaidacea Distribution
Family Parapseudidae Gutu, 1981
Genus Pseudohalmyrapseudes Larsen
& Hansknecht, 2004
P. aquadulcis Larsen & Hansknecht, 2004 ‘‘Freshwater spring’’ (but 1.93% in salinity), Australian Northern Territory
P. mussauensis (Shiino, 1965) ‘‘Brackish lake’’, Bismarck Archipelago (Papua New Guinea)
Family Tanaidae Dana, 1849
Genus Sinelobus Sieg, 1980
S. stanfordi (Richardson, 1901) Marine, plus freshwater, hypohaline and hypersaline inland waters of Galapagos,
Japan, Hong Kong, New Zealand, Australia, Argentina, Kurile Islands,
West Indies, Florida and Brazil (see Larsen & Hansknecht, 2004, and
references therein)
Family Anarthruridae Lang, 1971
Genus Paraleptognathia Kudinova-Pasternak,
1981
P. longiremis (Lilljeborg, 1864) Deep sea plus… Lake Baikal! (Kudinova-Pasternak, 1972) Record requiring
confirmation
228 Hydrobiologia (2008) 595:225–230
123

Larsen & Hansknecht (2004) suggest that Pseudohal-
myrapseudes occupies an intermediate position
between the euryhaline genus Halmyrapseudes Ba
˘
ce-
scu & Gutu, 1972 and Longiflagrum Gutu, 1995,
although no phylogenetic analysis was performed to
support this sugges tion. The Australian species is
inferred to have reached the spring it inhabits via the
groundwater system, although the possibility of an
upstream migration from the ocean cannot be ruled
out.
Two other species of tanaidaceans have been
reported from non-mar ine habitats, but both occur
also in marine environments. Sinelobus stanfordi
(Tanaidae), a widely distributed euryharine taxo n, has
been reported repeatedly from geographically scat-
tered freshwater, hypohaline or hypersaline lakes
(Tables 2 and 3; Fig. 1). In addition, there is a
doubtful record of the deep-sea trench Paraleptogna-
thia longiremis (Anarthruridae) from Lake Baikal
(Kudinova-Pasternak, 1972; Tables 2 and 3; Fig. 1).
This record requires confirmation and, as Larsen &
Hansknecht (2004) point out, the conspecificity of the
non-marine populations of these two taxa to their
corresponding marine forms should be confirmed,
suggesting that the current diversity of non-marine
tanaidacean species is underestimated.
Acknowledgement This is a contribution to Spanish MEC
project CGL2005-02217/ BOS.
References
Antsulevich, A. E., 2005. First finding of Cumacea crustaceans
in the Gulf of Finland. Vestnik Sankt-Peterburgskogo
Universiteta: Seriia 3—Biologiia 1: 84–87 (in Russian
with English summary).
Ba
˘
cescu, M., 1992. Cumacea II. Crustaceorum Catalogus 8:
175–468.
Ba
˘
cescu, M. & I. Petrescu, 1999. Ordre des Cumace
´
s (Cuma-
cea Krøyer, 1846). In Forest, J. (ed.), Traite
´
de Zoologie.
Anatomie, Syste
´
matique, Biologie. Tome VII (Fascicule
III A), Crustace
´
sPe
´
racarides. Me
´
moires de l’Institut Oc-
e
´
anographique, Monaco 19: 391–428.
Czerniavsky, V., 1868. Materialia ad zoographiam Ponticam
comparatum. Transactions of the first meeting of Russian
naturalists in Saint Petersburg, 1868: 19–136, 8 pls.
Derzhavin, A., 1912. Neue Cumaceen aus dem Kaspischen
Meer. Zoologischer Anzeiger 39: 273–284.
Dumont, H. J., 2000. Endemism in the Ponto-Caspian Fauna,
with special emphasis on the Onychopoda (Crustacea).
Advances in Ecological Research 31: 181–196.
Jones, N. S. & W. D. Burbanck, 1959. Almyracuma proximo-
culi gen. et spec. nov. (Crustacea, Cumacea) from
brackish water of Cape Cod, Massachusetts. Biological
Bulletin 116: 115–124.
Kudinova-Pasternak, R. K., 1972. Notes about the tanaidacean
fauna (Crustacea, Malacostraca) of the Kirmadec Trench.
Complex Research of the Nature of the Ocean. Publica-
tions Moscow University 3: 257–258.
Larsen, K. & T. Hansknecht, 2004. A new genus and species of
freshwater tanaidacean Pseudohalmyrapseudes aquadul-
cis (Apseudomorpha: Parapseudidae), from Northern
Territory, Australia. Journal of Crustacean Biology 24:
567–575.
Mordukhai-Boltovskoi, F. D. & N. N. Romanova, 1973. A new
genus of Cumacea from the Caspian Sea. Zoologicheskii
Zhurnal 52: 429–432.
Roccatagliata, D., 1981. Claudicuma platensis gen. et sp. nov.
(Crustacea, Cumacea) de la ribera argentina del Rı
´
odela
Plata. Physis (Buenos Aires) 39: 79–87.
Roccatagliata, D., 1991. Claudicuma platense Roccatagliata,
1981 (Cumacea): a new reproductive pattern. Journal of
Crustacean Biology 11: 113–122.
Sars, G. O., 1894. Crustacea Caspia, part II. Cumacea. Bulletin
de l’Academie Impe
´
riale des Sciences de St Petersburg
16: 297–338.
Sars, G. O., 1897. On some additional Crustacea from the
Caspian Sea. Annuaire du Muse
´
e Zoologique de
l’Acade
´
mie Imperiale des Sciences. St Petersburg 1897:
1–73, 8 pls.
Table 3 Global and per biogeographic region diversity (species number) of non-marine Cumacea and Tanaidacea
PA NA NT AT OL AU PAC World
Nannastacidae 1 (1) 1 (1) 2 (2)
Pseudocumatidae 19 (9) 19 (9)
Total Cumacea 19 (9) 1 (1) 1 (1) 21 (11)
Anarthruridae 1 (1) 1 (1)
Parapseudidae 2 (1) 2 (1)
Tanaidae 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1)
Total Tanaidacea 2 (2) 1 (1) 1 (1) 1 (1) 3 (2) 1 (1) 4 (3)
In brackets, number of genera. No records of these groups exist from Antarctica. PA: Palaearctic, NA: Nearctic, NT: Neotropical,
AT: Afrotropical, OL: Oriental, AU: Australasian, PAC: Pacific Oceanic Islands, ANT: Antarctic.
Hydrobiologia (2008) 595:225–230 229
123

Sars, G. O., 1914. Report on the Cumacea of the Caspian
Expedition 1904. Trudy KaspiıˇskoıˇE
´
kspeditzı¯ 1904
ghoda. 4: 1–34 (in Russian), 1–32 (in English), 12 pls.
Shiino, S. M., 1965. Tanaidacea from the Bismarck Archipel-
ago. Videnskabelige Meddelelser fra Dansk
Naturhistorisk Forening i Kjøbenhavn 128: 177–203.
Simpson, K. W., J. P. Fagnani, D. M. DeNicola & R. W. Bode,
1985. Widespread distribution of some estuarine
crustaceans (Cyathura polita, Chiridotea almyra, Almy-
racuma proximoculi) in the limnetic zone of the Lower
Hudson River, New York. Estuaries 8: 373–380.
Sowinski, W., 1893. Report on the Crustacea collected by Dr.
Ostroumow in the Sea of Azov. Zapiski Kievskago Ob-
shchestva Estestvoispytatelei 14: 289–405, (in Russian).
230 Hydrobiologia (2008) 595:225–230
123

FRESHWATER ANIMAL DIVERSITY ASSESSMENT
Global diversity of Isopod crustaceans (Crustacea; Isopoda)
in freshwater
George D. F. Wilson
Springer Science+Business Media B.V. 2007
Abstract The isopod crustaceans are diverse both
morphologically and in described species numbers.
Nearly 950 described species (*9% of all isopods)
live in continental waters, and possibly 1,400 species
remain undescribed. The high frequency of cryptic
species suggests that these figures are underestimates.
Several major freshwater taxa have ancient biogeo-
graphic patterns dating from the division of the
continents into Laurasia (Asellidae, Stenasellidae)
and Gondwana (Phreatoicidea, Protojaniridae and
Heterias). The suborder Asellota has the most
described freshwater species, mos tly in the families
Asellidae and Stenasellidae. The suborder Phrea to-
icidea has the largest number of endemic genera.
Other primary freshwater taxa have small numbers of
described species, although more species are being
discovered, especially in the southern hemisphere.
The Oniscidea, although primarily terrestrial, has a
small number of freshwater species. A diverse group
of more derived isopods, the ‘Flabellifera’ sensu lato
has regionally important species richness, such as in
the Amazon River. These taxa are transitional
between marine and freshwater realms and represent
multiple colonisations of continental habitats. Most
species of freshwater isopods species and many
genera are narrow range endemi cs. This endemism
ensures that human demand for fresh water will place
these isopods at an increasing risk of extinction, as
has already happened in a few documented cases.
Keywords Isopoda Crustacea Gondwana
Laurasia Diversity feeding Reproduction
Habits Fresh water Classification
Introduction
The Isopoda are a diverse group of crustaceans, with
more than 10,300 species found in all realms from the
deepest oceans to the montane terrestrial habitats;
approximately 9% of these species live in continental
waters. Isopods are thought of as dorsoventrally
flattened, and indeed many species fit this morpho-
logical stereotype. Diverse taxa found in the deep sea
and those found in groundwater habitats depart
considerably from this generalised body plan. Pala-
eontogical and phylogenetic evidence (Brusca &
Wilson, 1991; Wilson & Edgecombe, 2003) suggests
that the ancestral isopod may have had a narrow
Guest editors: E. V. Balian, C. Le
´
ve
ˆ
que, H. Segers &
K. Martens
Freshwater Animal Diversity Assessment
Electronic supplementary material The online version of
this article (doi:10.1007/s10750-007-9019-z) contains supple-
mentary material, which is available to authorized users.
G. D. F. Wilson (&)
Invertebrate Zoology, Australian Museum, 6 College
Street, Sydney, NSW 2010, Australia
e-mail: buz.wilson@austmus.gov.au
123
Hydrobiologia (2008) 595:231–240
DOI 10.1007/s10750-007-9019-z
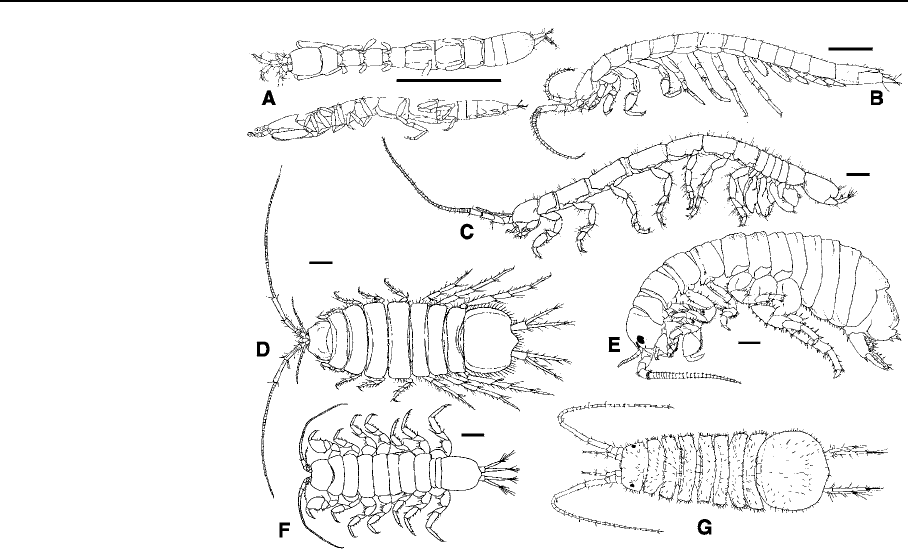
vaulted body with legs projecting ventrally (Fig. 1E,
Eophreatoicus, Amphisopidae). Freshwater taxa
include either typical flattened isopods (Fig. 1D, F,
G) or narrow body forms (Fig. 1B, C, E), along with
a few taxa that are thin and vermiform, often with
legs that emerge close to the dorsa l surface (Fig. 1A).
Other peculiarities of isopods include respiration
using their broad posterior limbs (swimming legs or
pleopods) with the heart positioned in the posterior
part of the body, and biphasic moulting, wherein the
back part of the body is cast off before the anterior
part. Limb forms are diverse in the isopods, but the
first walking leg (second thoracic limb) is modified
for grasping in most species.
Feeding
Isopods have a broad range of feeding types from
omnivory in Sphaeromatidae to carnivory in the
Cirolanidae. Oniscideans and Asellidae are well-
known as leaf litter shredders and have bacterial
endosymbionts to aid digestion (Zimmer, 2002,
Zimmer & Bartholme, 2003). Tainisopidae may be
carnivorous scavengers because they can be captured
using baited traps. Most freshwater isopods (e.g.
Asellota or Phreatoicidea) can be characterised as
generalised detritivores-omnivores, but may faculta-
tively choose other items. Phreatoicideans feed on
decaying vegetation and roots, or perhaps the micro-
flora and microfauna associated with these substrates
(Wilson & Fenwick, 1999), but on occasion will
engage in carnivory. Among the 942 described
species found in continental waters, the presumptive
feeding types (based on extrapolation from taxa
where habits are known) are as follows: 3.2% are
carnivores, 6.9% scavenger-carnivores, 9.9% ecto-
parasites, 0.4% herbivores, 6.1% omnivores and the
remaining 73.5% are detritivores-omnivores, mostly
Asellota and Phreatoicidea.
Reproduction
Isopods, like all peracarid crustaceans, have direct
development with the young brooded in a ventral
pouch until they are released as small adults. Isopods
have internal fertilisation (Wilson, 1991) that occurs
prior to the release of emb ryos into the marsupium,
unlike other peracarid crustaceans. Brood sizes range
from 4–5 young in tiny interstitial isopods to
hundreds in the parasitic forms, and lifetime
Fig. 1 Freshwater Isopoda,
a selection of body forms.
(A) Microcerberidae sp.
(interstitial, Western
Australia), dorsal and lateral
view; (B) Pygolabis sp.,
Tainisopidae (hypogean,
Western Australia); (C)
Phreatoicoides gracilis,
Hypsimetopidae (epigean,
Victoria Australia); ( D)
Asellus aquaticus, Asellidae
(epigean, Europe, from Sars
1897); (E ) Eophreatoicus
sp., Amphisopidae
(epigean, Northern
Territory Australia); (F),
Stenasellus chapmani,
Stenasellidae (hypogean,
Indonesia, from Coineau
et al. 1994; (G) Heterias
sp., Janiridae (hyporheic &
pholoteric, South America;
from Bowman et al. 1987).
Scale bars 1 mm, except for
A, 0.5 mm
232 Hydrobiologia (2008) 595:231–240
123

reproduction may be limited to one or several broods
in most species (Johnson et al., 2001). Many isopods,
especially the suborders Asellota and Oniscidea, have
secondary sexual features for intromission in both
males and females that are also useful for systemat-
ics. Brooding of the young, direct development and
internal fertilisation may be major contributory
factors in the high degree of endemism observed in
most isopod taxa (Wilson, 1991).
Habitats
Isopods occur in epigean lotic and lentic habitats (e.g.
Asellidae like the common European Asellus aquat-
icus and Phreatoicidae in Tasmania), but many live in
a variety of subterranean habitats. The Microcerberi-
dae are found interstitially in freshwater or marine
sands. Many families are limited to cavernicolous or
subterranean habitats, such as Stenasellidae, Microp-
arasellidae, or Tainisopidae. North American and
European members of the Asellidae can be both
epigean and hypogean (e.g. Turk et al., 1996; Lewis
& Bowman, 1981). Some taxa (e.g. Hypsimetopidae
or Heterias, Janiridae) could best be described as
infaunal, living in near subsurface habitats, either
burrowing among submerged roots, living in sub-
merged burrows of other animals (pholoteros) or in
the subsurface water of streams (hyporheos). A few
isopods occur in unusual habitats, such as Thermos-
phaeroma thermophilum in hot springs of the USA
southwest. Some oniscideans, which are ordinarily
terrestrial, have re-invaded the continental saline
waters (e.g. Haloniscus searlei) or even normal
freshwater (e.g. Trichoniscidae and Styloniscidae).
Australian collection records suggest that some
Philosciidae and Trichoniscidae may be amphibious
(see also Taiti & Humphreys, 2001; Tabacaru, 1999).
Methods
(See additional information on the article webpage).
The biodiversity of freshwater isopods is derived
from my research on the Phreatoicidea and Asellota,
and from the online ‘‘World List of Isopoda’’
(Kensley et al., 2005). The classification is derived
from that list (not as in Banerescu, 1990), but
includes the informal taxon ‘Flabellifera’ sensu lato
(see Wilson, 1999). The World List uses the tradi-
tional classification of the ‘Flabellifera’ that is known
to be paraphyletic (Brusca & Wilson, 1991; Wa
¨
gele,
1989; Tabacaru & Danielopol, 1999). The Microcer-
beridea includes two families, Microcerberidae and
Atlantasellidae (not Asellota as in Banarescu, 1990;
Wa
¨
gele, 1983; Jaume, 2001). The peculiar family
Calabozidae is classified as Oniscidea owing to its
possession of in-group genitalia and coxal plates
incorporated into the body (Brusca & Wilson, 1991).
Marine species, including those from anchialine cave
and marine beach interstitial environments, were
filtered out of the downloaded list, either using the
type habitat from the list or by consulting the original
literature. The data included species from saline
continental waters, such as Haloniscus. Subspecies
records were treated as species-level taxa. Unde-
scribed species (e.g. Heterias species) known to me
were added to the list where possible, although less
than 100 species were added. An estimate of the
unknown species was determined for the Phreatoici-
dea (Wilson in progress; see supplementary
information), and information from Gouws et al.
(2004, 2005). A diversity estimate for other isopod
groups used the simple known to unknown ratio from
the Phreatoicidea as applied to the other taxa
(Table 1). Although the assumption of similarity
between Phreatoicidea and other freshwater isopods
has obvious problems, this procedure at least pro-
vides an hypothesis for further refinement.
Species diversity
Of the entirely freshwat er isopods (marked with an
asterisk in Table 1), the Asellota has the most of the
942 described species, with the largest number of
species in the family Asellidae, followed by the
Stenasellidae. The Phreatoicidea have at least four
families with many undescribed species (see Table in
supplementary information) that may double the
number of described species. Other freshwater fam-
ilies have small numbers of described species,
although more species are being discovered as
surveys are carried out in the southern hemisphere.
The Protojaniridae are tiny and fragile, and may
require specialis ed techniques to recover them from
hypogean habitats; 12 species in five genera are
described, but more remain to be found. Recently,
Hydrobiologia (2008) 595:231–240 233
123

J. Pe
´
rez-Schultheiss in Chile sent specimens of a new
protojanirid; anot her new species is known from
northern Australia. The application of ‘‘known to
unknown’’ estimates from the Phrea toicidea to the
other freshwater isopods results in 62% more than
those known, or approximately a total of 2,630
species (Table 1).
Evidence from molecular studies suggest that this
estimate could be highly conservative. RAPD (ran-
dom amplified polymorphic DNA) studies on both
species of Asellidae and Stenasellidae (Baratti et al.,
1999; Verovnik et al., 2003) have uncovered previ-
ously unsusp ected diversity in well-known
populations of Stenasellus and Proasellus. Similar
results have been obtained from studies of genetic
variation using enzymatic loci (Proasellus: Ketmaier,
2002) or the mtDNA cytochrome oxidase I gene (CO-
I) (Stenasellus: Ketmaier et al., 2003). Cryptic spe-
cies in the epigean phreatoicidean genus
Mesamphisopus (Gouws et al., 2004, 2005) could
Table 1 Species Diversity of Freshwater Isopoda. Estimation method and classification explained in text and supplementary
material (see additional information)
Suborder Family Species, described
and new
Estimated
unknown species
Estimated
total diversity
PHREATOICIDEA
Stebbing, 1893
*
Amphisopidae Nicholls, 1943 36 48 84
*
Hypsimetopidae Nicholls, 1943 11 19 30
*
Incertae sedis (Crenisopus)1 1
*
Phreatoicidae Chilton, 1891 49 71 120
*
Ponderellidae Wilson & Keable, 2004 2 2
Subtotal, used for estimates other suborders 99 138 237
Unknown to Known ratio 1.39
ASELLOTA Latreille,
1803
*
Asellidae Rafinesque-Schmaltz, 1815 379 529 908
Janiridae G. O. Sars, 1897 76 106 182
Microparasellidae Karaman, 1933 73 102 175
*
Protojaniridae Fresi, Idato & Scipione, 1980 15 21 36
*
Stenasellidae Dudich, 1924 73 102 175
MICROCERBERIDEA
Lang, 1961
Microcerberidae Karaman, 1933 21 30 51
ONISCIDEA Latreille,
1803
*
Calabozoidae Van Lieshout, 1983 2 3 5
Philosciidae Kinahan, 1857 1 2 3
Scyphacidae Dana, 1852 5 7 12
Trichoniscidae Sars, 1899 1 2 3
‘FLABELLIFERA’
sensu lato
Aegidae Leach, 1815 1 2 3
Anthuridae Leach, 1814 19 27 46
Bopyridae Rafinesque-Schmaltz, 1815 33 46 79
Cirolanidae Dana, 1852 65 91 156
Corallanidae Hansen, 1890 4 6 10
Cymothoidae Leach, 1818 51 72 123
Entoniscidae Kossmann, 1881 4 6 10
Idoteidae Samouelle, 1819 4 6 10
Leptanthuridae Poore, 2001 3 5 8
Paranthuridae Menzies & Glynn, 1968 1 2 3
Sphaeromatidae Latreille, 1825 57 80 137
*
Tainisopidae Wilson, 2003 7 10 17
Total 994 1395 2625
(*Entirely freshwater families)
234 Hydrobiologia (2008) 595:231–240
123