Balian E.V., L?v?que C., Segers H., Martens K. (Eds.) Freshwater Animal Diversity Assessment
Подождите немного. Документ загружается.

account for a large percentage of mysid species
diversity.
Tethyan subterranean relicts
All of the subterranean/groundwater mysid genera
considered here (Antromysis ––6 spp., Spelaeomysis––
7 spp., Stygiomysis––5 spp., and the monotypic
Troglomysis vjetrenicensis) are found in a
distributional pattern suggesting a Tethy an origin,
likely colo nizing groundwater habitats due to the uplift
and stranding of marine ancestors during Miocene
regressions of the Tethys and Mediterranean seas
(Boxshall & Jaume, 2000). These hypothesized Tethy-
ian dispersal events resulted in ancient mysid genera
successfully colonizing early regions of Central Amer-
ica, the Caribbean, and Indian and Mediterranean
basins, resulting in widespread genera that were later
isolated, and presently represented by species often
Table 2 Total number of inland mysid genera from the major
geographical regions from each family (**), subfamily (*), and
tribe () containing at least one inland genus. Only genera from
each family are used to calculate the final totals for each
region. See supplementary tables for detailed information. PA:
Palaearctic; NA: Nearctic; NT: Neotropical; AT: Afrotropical;
OL: Oriental; AU: Australasian; PAC: Pacific & Oceanic
Islands; ANT: Antarctic
PA NA NT AT OL AU PAC ANT Total inland genera
**Lepidomysidae 1 1 10100 0 1
**Stygiomysidae 1 0 10000 0 1
**Mysidae 13 6 41510 0 23
*Rhopalophthalminae 0 0 00100 0 1
*Mysinae 13 6 41410 0 22
Heteromysini 0 1 0000 0 0 1
Leptomysini 0 1 10010 0 2
Mysini 13 4 31400 0 19
Total 15 7 61610 0 25
Note: Several genera occur in more than one geographical region, resulting in higher numbers of occurrences (36) than inland genera
(25)
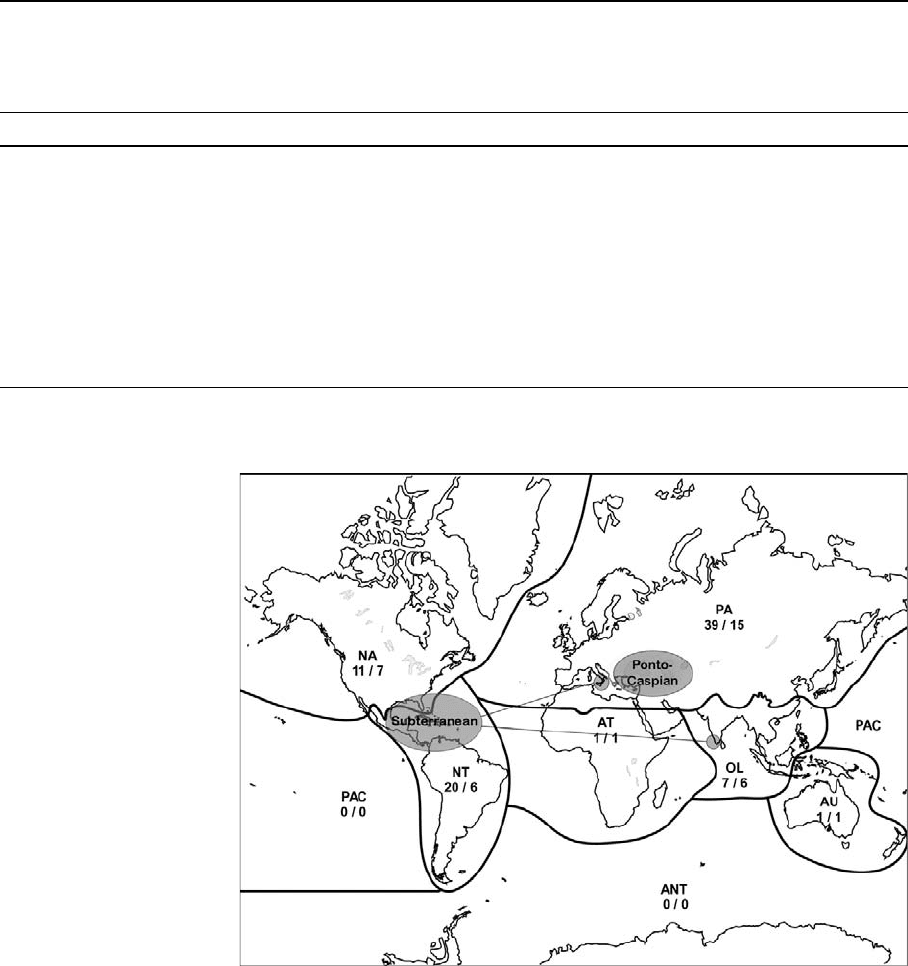
Fig. 2 Biogeographic
regions indicating the
numbers of inland mysid
species and genera (SP/GN)
found in each: PA––
Palaearctic; NA––Nearctic;
NT––Neotropical; AT––
Afrotropical; OL––Oriental;
AU––Australasian; PAC––
Pacific; ANT––Antarctica;
Grey circles indicate areas
of high biodiversity, i.e., the
Ponto-Caspian region and
the distribution of
Lepidopmysidae and
Stygiomysidae continental
subterranean species (lines
connect disjunct regions of
occurrence)
Hydrobiologia (2008) 595:213–218 215
123

endemic to a single groundwater system. Additionally,
the surface genera Parvimysis (2 spp.) and Surinamysis
(3 spp.), distributed in the Ca ribbean and South
America, are closely related to the genus Antromysis
and may also be a part of this Tethyan distribution.
Autochthonous Ponto-Caspian endemics
The Ponto-Caspian basin, consisting of the Black,
Azov, and Caspian Seas, is composed of inland seas
with complex geological histories dating back to the
Paratethys Sea (20 Mya), including periods as lacus-
trine environments (Banarescu, 1991). The Ponto-
Caspian mysid fauna (autochthonous + ‘glacial re-
licts’, see below) are generally considered the center
of inland mysid species diversity. The autochthonous
mysids that evolved in these enclosed continental
basins, occur in fresh and brackish water portions of
the basins includi ng rivers, lakes, and estuaries, and
are endemic to one basin or are found in parts of all
three (Table 3). Although this fauna consists of seven
genera and 20 species, a large portion of the endemic
mysid dive rsity in these basins is the result of a
radiation in the genus Paramysis (Table 3).
In a phylogeographic study of Limnomysis bene-
deni and six Paramysis species across the Ponto-
caspian region (Audzijonyte
¨
et al., 2006), three main
patterns were identified: (1) no deep subdivisions
across the entire region, (2) genealogical splits
matching geographical borders among basins, and
(3) divergent lineages occurring only within the
Caspian. The discordant molecular subdivisions
among these co-distributed mysid species suggest
that the similar zoogeographic patterns were formed
at different times (Audzijonyte
¨
et al., 2006).
Mysis ‘glacial relicts’
The genus Mys is comprises 14 species, of which
eight have inland freshwater distributions and can be
divided into two groups: (1) The M. relicta group (M.
relicta, M. diluviana, M. salemaai, M. segerstralei)
with a circumpolar distribution from boreal and
subarctic lakes of the previously glaciated areas of
Europe and North America (Audzijonyte
¨
&Va
¨
ino
¨
la
¨
,
2005), and (2) four Caspian Sea endemics (Table 3).
Phylogenetic analyses indicate that the inland Mysis
species are a monophyletic assemblage, sister to
circumarctic marine species (Audzijonyte
¨
et al.,
2005). The separation of freshwater/continental Mysis
spp. and circumarctic marine species is estimated to
have taken place 3–7 Mya (Audzijonyte
¨
et al., 2005),
and does not correspond to the timing in general
hypotheses of continental invasions, such as mid-
Pleistocene glaciation events or mid-Tertiary separa-
tion of the Arctic and Caspian basins (see Va
¨
ino
¨
la
¨
,
1995). Furthermore, molecular divergences among
the boreal Nearctic (M. diluviana) and Palearctic (M.
relicta, M. salemaai, M. segerstralei) species and the
Caspian endemics indicate inland colonizations
Table 3 List of species endemic to the Black, Azov, and
Caspian Seas, including the occurrence in each basin (data from
Audzijonyte
¨
, 2006). In addition to the species counted here,
there are at least four additional endemic Ponto-Caspian mysids
(Diamysis mecznikovi, Hemimysis serrata, Paramysis agigen-
sis, Paramysis pontica) whose distributions are unclear or do
not occur in freshwaters and therefore have not been included
in the species list
Black Sea Azov Sea Caspian Sea
‘Glacial-relict’
Mysis amblyops ·
Mysis caspia ·
Mysis macrolepis ·
Mysis microphthalma ·
Autochtonous Ponto-Caspian
Caspiomysis knipowitschi ·
Diamysis pengoi ·
Diamysis pusilla ·
Hemimysis anomala ···
Katamysis warpachowsky ···
Limnomysis benedeni ···
Paramysis baeri ···
Paramysis eurylepis ·
Paramysis grimmi ·
Paramysis incerta ·
Paramysis inflata ·
Paramysis intermedia ···
Paramysis kessleri ··
Paramysis kosswigi ·
Paramysis kroyeri ·
Paramysis lacustris ···
Paramysis loxolepis ·
Paramysis sowinskii ··
Paramysis ullskyi ···
Schistomysis elegans ·
216 Hydrobiologia (2008) 595:213–218
123

occurring at different times (A udzijonyte
¨
&Va
¨
ino
¨
la
¨
,
2006). M. relicta and M. diluviana are considered the
oldest of Mysis freshwater species that independently
colonized their respective European and American
ranges during early Pleistocene (Va
¨
ino
¨
la
¨
et al.,
1994). In contrast, M. salemaai and M. segerstralei
are younger closely related species that have pene-
trated freshwaters more recently (Audzijonyte
¨
&
Va
¨
ino
¨
la
¨
, 2006). As for the four endemic Caspian
Mysis species, small molecular divergences suggest a
recent, possibly late Pleistocene, sympatric radiation,
possibly driven by adaptation to a deep pelagic
habitat by M. amblyops and M. microphthalma
(Va
¨
ino
¨
la
¨
, 1995; Audzijonyte
¨
et al., 2005).
Euryhaline estuarine fauna
Most of the remaining mys ids (20 spp., 14 genera) are
euryhaline species with at least one population
occurring in marginal freshwaters. These species
have only very recently invaded freshwaters in
portions of their distributions.
Phylogeny
The taxonomic position and phylogenetic affiliations
within the Mysida are currently under debate.
Historically, the Mysida were considered members
of the crustacean superorder Peracarida, placed as a
sister taxon to the Lophogastrida in the order
Mysidacea. More recently, molecular studies have
led to raising the Mysida (and Lophogastrida) to
ordinal rank (Spears et al., 2005). With respect to
those families containing inland fauna, there is also
taxonomic and phylogenetic uncertainty. For exam-
ple, molecular and morphological data show that the
subterranean family Stygiomysidae is more closely
related to the order Mictacea than to other Mysida
families, suggesting that they be removed from the
order Mysida and placed within a separate order,
Stygiomysida, comprising the families Stygiomysi-
dae and Lepidomysidae (Meland & Willassen, 2007).
Conservation issues
At least 19 inland mysid species are associated with
groundwater habitats (caves, wells, and crab burrows)
having very limited areas of distribution that are
highly susceptible to pollution from the surface.
These species in particular are in need of assessment
for conservation ranking, as they are often found in
aquifers important to local communities as a source
of freshwater and may serve as indicators of water
quality. Many freshwater mysid species have also
served an important role in both US and European
fisheries, where they have been introduced into lakes
and reservoirs to serve as food for commercially
important fish species (Mordukhai-Boltovskoi, 1979;
Northcote, 1991).
In contrast, the autochthonous Ponto-Caspian
mysids are currently invading aquatic ecosystems of
Northern Europe as a result of human activities (De
Vaate et al., 2002; Leppakoski et al., 2002). The
impact of invasive mysid species on native lacustrine
and riverine ecosystems can be large, including a
severe reduction in zooplankton abundance, with
concomitant negative effects on higher consumers
(Spencer et al., 1991; Ketelaars et al., 1999).
Acknowledgments We thank R. Va
¨
ino
¨
la
¨
for advice
concerning the Ponto-Caspian mysid fauna and for the
helpful suggestions of one anonymous reviewer. T. bowmani
and A. almyra images by E. Peebles from ‘Common and
Scientific Names of Aquatic Invertebrates from the United
States and Canada: Crustaceans’ are courtesy of the American
Fisheries Society. This work was supported by NSF grant
DEB-0206537.
References
Audzijonyte
¨
, A., 2006. Diversity and zoogeography of conti-
nental mysid crustaceans. Walter and Andre
´
e de Nottbeck
Foundation Scientific Reports 28: 1–46.
Audzijonyte
¨
, A. & R. Va
¨
ino
¨
la
¨
, 2005. Diversity and distribu-
tions of circumpolar fresh- and brackish-water Mysis
(Crustacea: Mysida): descriptions of M. relicta Love
´
n,
1862, M. salemaai n. sp., M. segerstralei n. sp. and M.
diluviana n. sp., based on molecular and morphological
characters. Hydrobiologia 544: 89–141.
Audzijonyte
¨
, A., & R. Va
¨
ino
¨
la
¨
, 2006. Phylogeographic anal-
yses of a circumarctic coastal and a boreal lacustrine
mysid crustacean, and evidence of fast postglacial
mtDNA rates. Molecular Ecology 15: 3287–3301.
Audzijonyte
¨
, A., M. E. Daneliya, & R. Va
¨
ino
¨
la
¨
, 2006. Com-
parative phylogeography of Ponto-Caspian mysid
crustaceans: isolation and exchange among dynamic
inland sea basins. Molecular Ecology 15: 2969–2984.
Audzijonyte
¨
, A., J. Damgaard, S.-L. Varvio, J. K. Vainio & R.
Va
¨
ino
¨
la
¨
, 2005. Phylogeny of Mysis (Crustacea, Mysida):
history of continental invasions inferred from molecular
and morphological data. Cladistics 21: 575–596.
Hydrobiologia (2008) 595:213–218 217
123

Banarescu P., 1991. Zoogeography of Fresh Waters, Vol. 2.
Distribution and Dispersal of Freshwater Animals in
North American and Eurasia. Aula Verlag, Wiesbaden.
Boxshall, G. A. & D. Jaume, 2000. Discoveries of cave
misophrioids (Crustacea: Copepoda) shed new light on the
origin of anchialine faunas. Zoologischer Anzeiger 239:
1–19.
De Vaate, A. B., K. Jazdzewski, H. A. M. Ketelaars, S. Gol-
lasch, G. Van der Velde, 2002. Geographical patterns in
range extension of Ponto-Caspian macroinvertebrate
species in Europe. Canadian Journal of Fisheries and
Aquatic Sciences 59: 1159–1174.
Ketelaars, H. A. M., F. E. Lambregts-van de Clundert, C. J.
Carpentier, A. J. Wagenvoort & W. Hoogenboezem,
1999. Ecological effects of the mass occurrence of the
Ponto-Caspian invader, Hemimysis anomala GO Sars,
1907 (Crustacea: Mysidacea), in a freshwater storage
reservoir in the Netherlands, with notes on its autecology
and new records. Hydrobiologia 394: 233–248.
Leppakoski, E., S. Golllasch, P. Gruszka, H. Ojaveer, S. Olenin
& V. Panov, 2002. The Baltic: a sea of invaders. Canadian
Journal of Fisheries and Aquatic Science 59: 1175–1188.
Martin, J. W. & G. E. Davis, 2001. An updated classification of
the recent crustacea. Natural History Museum of Los
Angeles County, Science Series 39: 1–124.
Mauchline, J., 1980. The biology of mysids and euphausiids.
Advances in Marine Biology 18: 1–680.
Meland, K. & E. Willassen, 2007. The disunity of ‘‘Mysida-
cea’’ (Crustacea). Molecular Phylogenetics and Evolution.
http://dx.doi.org/10.1016/j.ympev.2007.02.009
Mordukhai-Boltovskoi, F. D., 1979. Composition and distri-
bution of Caspian fauna in the light of modern data.
Internationale Revue der Gesamten Hydrobiologie 64: 1–
38.
Northcote, T. G., 1991. Success, problems, and control of
introduced mysid populations in lakes and reservoirs.
American Fisheries Society Symposium 9: 5–16.
Spears, T., R. W. DeBry, L. G. Abele & K. Chodyla, 2005.
Peracarid monophyly and interordinal phylogeny inferred
from nuclear small-subunit ribosomal DNA sequences
(Crustacea: Malacostraca: Peracarida). Proceedings of the
Biological Society of Washington 118: 117–157.
Spencer, C. N., B. R. McClelland & J. A. Stanford, 1991.
Shrimp stocking, salmon collapse, and eagle displace-
ment. Bioscience 41: 14–21.
Va
¨
ino
¨
la
¨
, R., 1995. Origin and recent endemic divergence of a
Caspian Mysis species flock with affinities to the ‘‘glacial
relict’’ crustaceans in boreal lakes. Evolution 49: 1215–
1223.
Va
¨
ino
¨
la
¨
, R., B. J. Riddoch, R. D. Ward & R. I. Jones, 1994.
Genetic zoogeography of the Mysis relicta species group
(Crustacea: Mysidacea) in northern Europe and North
America. Canadian Journal of Fisheries and Aquatic
Science 51: 1490–1505.
Wittmann K. J., 1999. Global biodiversity in Mysidacea, with
notes on the effects of human impact. In Schram F. R. & J.
C. von Vaupel Klein (eds), Crustaceans and the Biodi-
versity Crisis. Proceedings of the Fourth International
Crustacean Congress, Amsterdam, The Netherlands, July
20–24, 1998, Vol. I. Brill NV, Leiden: 511–525.
218 Hydrobiologia (2008) 595:213–218
123

FRESHWATER ANIMAL DIVERSITY ASSESSMENT
Global diversity of spelaeogriphaceans
& thermosbaenaceans (Crustacea; Spelaeogriphacea
& Thermosbaenacea) in freshwater
D. Jaume
Springer Science+Business Media B.V. 2007
Abstract Spelaeogriphaceans and thermosbaena-
ceans are two orders of eyeless, unpigmented perac-
arid crustaceans represented by very few species from
subterranean waters. Spelaeogriphaceans occur only
in continental waters, either running or still, in
limestone or sandstone caves, or in calcrete aquifers.
The four species known are limnic except one
occurring in slightly brackish water loosely associ-
ated with an endorheic basin. The Thermosbaenacea
are primarily marine, with only 18 species recorded
in limnic conditions or in brackish inland waters
whose salinity does not derive from dilution of
seawater. They occur in limestone caves, the inter-
stitial medium associated to alluvial deposits, or in
thermo-mineral springs. Spelaeogriphaceans are
found on the southern continents, in ancient cratons
not affected by sea transgressions at least since the
Early Cretaceous, when Gondwana started to break-
up. The former integration of these terranes into
Gondwana suggests that the penetration of spelaeo-
griphaceans in continental waters took place previous
to the fragmentation of this super continent (starting
ca. 140 Ma), and that their current distribution
pattern was driven by continental drift. The distribu-
tion of the Thermosbaenacea matches precisely the
area covered by the ancient Tethys Sea or its
coastlines. They are most probably relicts of a once
widespread shallow-water marine Tethyan fauna
stranded in interstitial or crevicular groundwater
during marine regressions.
Keywords Freshwater Global assessment
Species richness Peracarida Crustacea
Introduction
Spelaeogriphaceans and thermosbaenaceans are two
orders of eyeless, unpigmented peracarid crustaceans
represented by very few species from subterranean
waters. The Spelaeogriphacea Gordon, 1957 occur
only in continental waters, either running or still, in
limestone or sandstone caves, or in calcrete aquifers.
All species are limnic except one occurring in slightly
brackish water loosely associated to an endorheic
basin.
Thermosbaenacea Monod, 1927, in contrast, seem
to be primarily marine although only 5 out of the 34
species known are euhaline, living in lava tubes or in
Guest editors: E. V. Balian, C. Le
´
ve
ˆ
que, H. Segers &
K. Martens
Freshwater Animal Diversity Assessment
Electronic supplementary material The online version of
this article (doi:10.1007/s10750-007-9017-1) contains
supplementary material, which is available to authorized
users.
D. Jaume (&)
IMEDEA (CSIC-UIB), Instituto Mediterra
´
neo de Estudios
Avanzados, c/Miquel Marque
`
s 21, Esporles, Illes Balears
07190, Spain
e-mail: d.jaume@uib.es
123
Hydrobiologia (2008) 595:219–224
DOI 10.1007/s10750-007-9017-1
the interstices between submersed coarse sand grains,
volcanic tephra or coral rubble (Wagner, 1994). Most
taxa occur in the anchialine environment associa ted
with marine coastal areas, where water salinity varies
sharply across the water column and derives from
dilution of sea water. Only 18 species have been
recorded in limnic conditions or in brac kish inland
waters whose salinity does not derive from dilution of
seawater. They occur in limestone caves, the inter-
stitial medium associated to alluvial deposits or in
thermo-mineral springs.
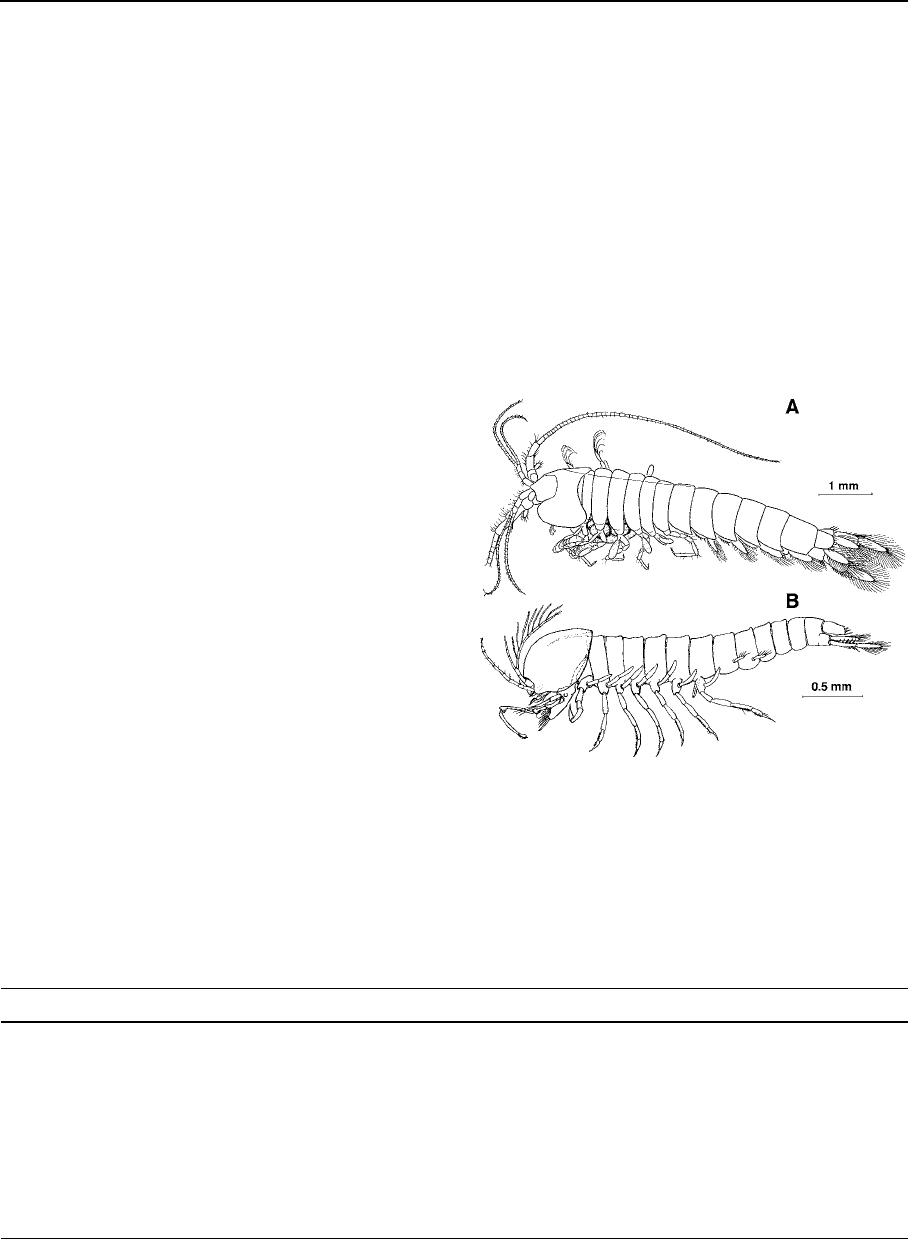
The spelaeogriphacean body is roughly cylindri-
cal, with the cephalothorax incorporating only the
first thoracomere (which carries the maxillipeds)
and with a short, posteriorly directed carapace cover-
ing only part of the second thoracomere dorsally
(Fig. 1A). The pereion consists of seven free pereio-
meres, each with a pair of similar biramous stenop-
odial pereiopods, and the pleon comprises six free
pleomeres, each with a pair of well-developed
biramous pleopods. The telson is free, articulated to
the 6th pleomere. The most remarkable autapom or-
phy of the group is the transformation of the exopods
of the posterior pairs of pereiopods into non-setose,
respiratory paddles.
Contrary to spelaeogriphaceans, whose inclusion
in the Peracarida is undeniable since brooding
females display a thoracic ventral marsupium formed
by oo
¨
stegites (= foliaceous medial exte nsions of the
pereiopodal coxae), the inclusion of the Thermos-
baenacea in this group is debatable since here the
embryos are carried in a dorsal brood pouch derived
from the carapace (Richter & Schol tz, 2001). Apart
of this, the rest of features of the therm osbaenacean
body plan is roughly similar to the spelaeogripha-
ceans (Fig. 1B); just notice that the pleopods are now
vestigial and present only on pleomeres 1 and 2, and
that the condition of the posterior extension of the
carapace varies from covering the second thoraco-
mere only (in males and non-brooding females of
Thermosbaenidae, Monodellidae and Halosbaenidae)
to cover all thoracomeres (Tulumellidae). Thermos-
baena mirabilis is a highly modified therm osbaena-
cean that separates from the ordinary morphology of
the group in having a pleotelson (formed by the 6th
pleomere and telson), and the display of only five
pairs of pereiopods.
The mouthparts of spelaeogriphaceans and ther-
mosbaenaceans conform to a functional series of
scrapers in an arrangement considered to fit for
Fig. 1 General aspect of Spelaeogriphacea and Thermosbaen-
acea. (A) Spelaeogriphus lepidops Gordon 1957, a spelaeogri-
phacean from Table Mountain, South Africa (after Gordon,
1960). (B) male of Tethysbaena atlantomaroccana (Boutin &
Cals, 1985), a freshwater thermosbaenacean from Morocco;
notice that ornamentation of pereopodal exopods is omitted
from figure (after Cals & Boutin, 1985)
Table 1 Global diversity of Spelaeogriphacea
Distribution Habitat Salinity range
Spelaeogriphidae
Mangkurtu Poore & Humphreys, 1998
mityula Poore & Humphreys, 1998 NW Australia Borehole wells Limnic
kutjarra Poore & Humphreys, 2003 NW Australia Borehole wells Oligohaline
Potiicoara Pires, 1987
brasiliensis Pires, 1987 Brazil Caves Limnic
Spelaeogriphus Gordon, 1957
lepidops Gordon, 1957 South Africa Caves Limnic
220 Hydrobiologia (2008) 595:219–224
123

scraping small food particles from a substrate (Fryer,
1964).
Species diversity
The Spelaeogriphacea comprises a single family, the
Spelaeogriphidae Gordon, 1957 with three genera:
Speleaeogriphus and Potiicoara (both monotypic),
and Mangkurtu, with two species (Table 1). Three
fossil forms from the Carboniferous of Canada, the
Upper Jurassic of China, and the Lower Cretaceous
of Spain are treated also as spelaeogriphaceans by
some authors (Schram, 1974; Shen et al., 1998;
1999). Nevertheless, none of these fossil forms
preserves the diagnostic pereopodal exopods, and
their body tagmosis and short carapace do not
preclude their allocation to other peracaridan orders.
The Thermosbaenacea embraces four families, of
which only three include non-marine species
(Table 2). The family Thermosbaenidae is mono-
typic. The Monodellidae includes two genera:
Table 2 Global diversity of non-marine Thermosbaenacea
Distribution Habitat Salinity range
Thermosbaenidae
Thermosbaena Monod, 1924
mirabilis Monod, 1924 Tunisia Thermo-mineral springs Presumed oligohaline
Monodellidae
Tethysbaena Wagner, 1994
juriaani Wagner, 1994 Dominican Rep. Anchialine limestone wells
and spring
Limnic to oligohaline
gaweini Wagner, 1994 Dominican Rep. Wells in alluvial sediments Limnic to oligohaline
haitiensis Wagner, 1994 Haiti Spring, wells and alluvial
sediments
Limnic to oligohaline
juglandis Wagner, 1994 Haiti Wells in alluvial sediments Limnic
lazarei Wagner, 1994 Cuba Cave and river interstitial Limnic to oligohaline
tinima Wagner, 1994 Cuba Wells in limestone Limnic
calsi Wagner, 1994 Saint John; Tortola (British Virgin
Islands)
Wells in alluvial deposits Limnic to oligohaline
relicta (Po
´
r, 1962) Israel Thermo-mineral springs Oligohaline to hyperhaline
somala (Chelazzi & Messana,
1982)
Somalia Wells in limestone Oligohaline to polyhaline
(evaporites)
atlantomaroccana (Cals &
Boutin, 1885)
Morocco Wells in alluvial deposits Presumed limnic
tarsiensis Wagner, 1994 Spain Well in alluvial deposit Limnic
texana (Maguire, 1965) Texas (U.S.A.) Artesian wells Limnic
vinabayesi Wagner, 1994 Isla Juventud (Cuba) Cave Limnic
Halosbaenidae
Limnosbaena Stock, 1976
finki (Mestrov & Lattinger-
Penko, 1969)
Bosnia-Hercegovina; Italy Interstitial of river alluvia;
caves
Limnic
sp. Wagner, 1994 France Well Limnic
Halosbaena Stock, 1976
tulki Poore & Humphreys, 1992 NW Australia Calcrete aquifers Oligohaline (evaporites)
Theosbaena Cals & Boutin,
1985
cambodjana Cals & Boutin,
1985
Cambodia; Thailand Caves Limnic
Salinity tolerance ranges: Limnic: <0.5%; Oligohaline: 0.5–5%; Polyhaline: 18–30%; Hyperhaline: >40%
Hydrobiologia (2008) 595:219–224 221
123
Monodella (monotypic) and Tethysbaena (23 spe-
cies), of which only 13 species of Tethysbaena are
considered here as non-marine. The Halosbaenidae
consists of the truly limnic genera Limnosbaena (two
species, one of them not formally described) and
Theosbaena (monotypic), plus the genus Halosbaena,
with two marine species plus one from non-marine,
brackish inland waters. The fourth thermosbaen acean
family Tulumellidae, comprising the single genus
Tulumella (three species), is fully marine. There is
no fossil record of the Thermosbaenacea known to
date.
Many species of thermosbaenaceans have been
reported from oligohaline water in coastal aquifers or
anchialine environments only, frequently around
haloclines where water salinity changes abruptly
from limnic-oligohaline to marine euhaline. These
taxa (shown in Appendix Table 1) are not included in
the total estimate of non-marine species since none
has been reported from pure fresh waters, nor from
marine euhaline water either.
Present distribution and historical processes
Living spelaeogriphaceans appear associated with
freshwater in southern continents, in ancient cratons
not affected by sea transgressions at least since the
Early Cretaceous, when Gondwana started to break-
up (Fig. 2; Table 3; Appendix Fig. A). Spelaeogri-
phus lepidops is known only from two caves in South
Africa excavated in Ordovician quartzites (Gordon,
1957). Potiicoara brasiliensis, from two caves in
Upper Proterozoic limestone at Mato Grosso do Sul
(Brazil; Pires, 1987). And the two species of Mang-
kurtu, from borehole wells in calcrete of Middle to
Late Tertiary age on north-western Australia,
although these deposits overlie and are in direct
contact with Early Cretaceous alluvial conglomerates
(Poore & Humphreys, 1998; 2003). The former
integration of these terranes in Gondwana suggests
that the penetration of spelaeogriphaceans in conti-
nental waters took place previous to the fragmenta-
tion of this supercont inent (starting ca. 140 Ma), and
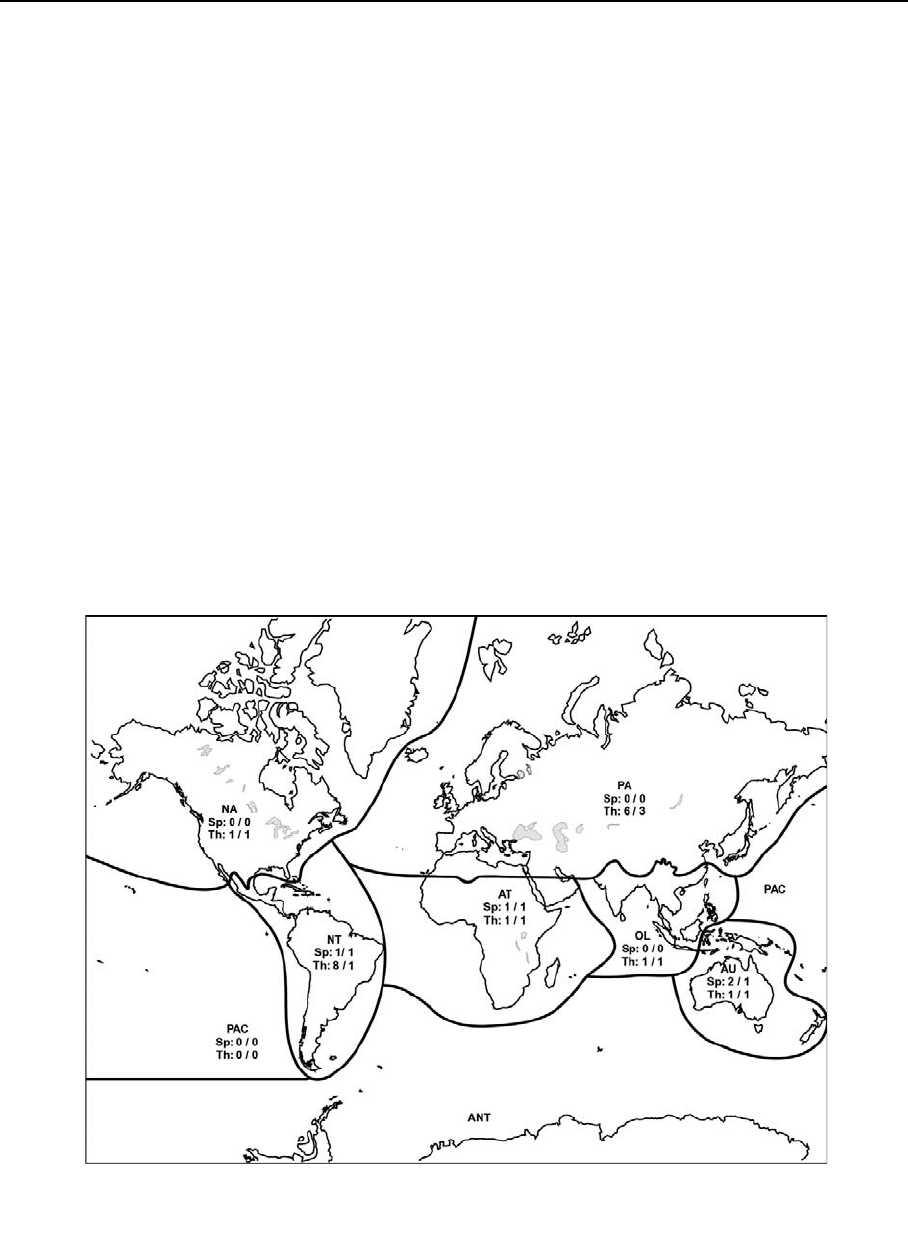
Fig. 2 Global distribution of Spelaeogriphacea and continen-
tal water Thermosbaenacea (Species number/Genus number).
PA––Palaearctic, NA––Nearctic, NT––Neotropical, AT––
Afrotropical, OL––Oriental, AU––Australasian, PAC––Pacific
Oceanc Islands, ANT––Antarctic
222 Hydrobiologia (2008) 595:219–224
123
that their current distribution pattern was driven by
continental drift.
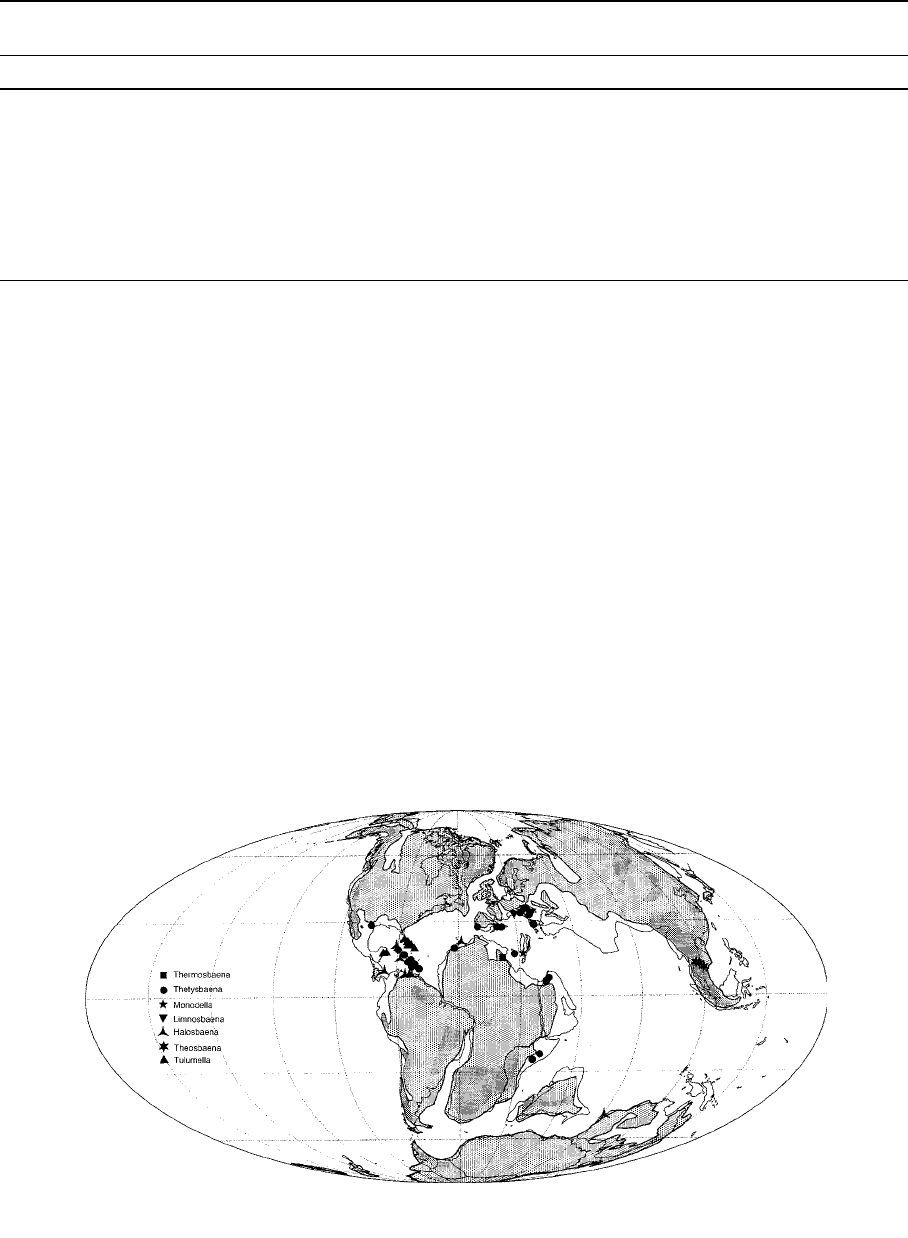
The distribution of the order Thermosbaenacea
matches precisely the area covered by the ancient
Tethys Sea or its coastlines (Fig. 2, 3; Table 3; see
also Appendix Figs A and B). They are most
probably relic ts of a once widespread shallow-water
marine Tethyan fauna, stranded in interstitial or
crevicular groundwater during episodes of marine
regression. The amphi-Atlantic distribution of Teth-
ysbaena, or the so-called ‘‘full Tethyan track’’
displayed by Halosbaena (with species in the Carib-
bean, Canary Islands and Australia) suggest the
origin of these genera dates back to at lea st the final
opening of the Atlantic (95 Ma) and the maximum
extent of the Tethys sea (120 Ma), respectively. The
timing of colonisation of continental waters by
thermosbaenacean lineages is probably more recent
(all inland water stations fall within areas covered by
the sea in Pliocene or more recent times), and the
speciation process can have been peripatric.
The distribution of the Thermosbaenidae casts
doubts on whether they represent a separate family
from its closest relative, the Monodellidae. The
single representative of the family dwells in a
Tunisian thermo-mineral spring placed in a zone
recently covered by the sea (Late Pliocene). This
station falls within the area covered by the Monod-
ellidae, which comprises a widespread amphi-Atlan-
tic genus (indicative of the ancient origin for the
family; see above) present also in N Africa (Teth-
ysbaena), plus a second genus Monodella, mono-
typic and kn ow only from a single-anchialine cave in
Italy. The Thermosbaenidae are probably a young,
Fig. 3 Translocation of present distribution of Thermosbaenacea (including both marine and continental water taxa) to an Aptian
(Lower Cretaceous; 120 Ma) palaeo-coastline map. Shaded areas denote emerged lands
Table 3 Global and per biogeographic region diversity (species number) of Spelaeogriphacea and non-marine Thermosbaenacea
PA NA NT AT OL AU PAC World
Spelaeogriphacea
Spelaeogriphidae – – 1 (1) 1 (1) – 2 (1) – 4 (3)
Thermosbaenacea
Thermosbaenidae 1 (1) – – – – – – 1 (1)
Monodellidae 3 (1) 1 (1) 8 (1) 1 (1) – – – 13 (1)
Halosbaenidae 2 (1) – – – 1 (1) 1 (1) – 4 (3)
Total Thermosbaenacea 6 (3) 1 (1) 8 (1) 1 (1) 1 (1) 1 (1) – 18 (5)
In brackets, number of genera. No records of these groups exist from Pacific oceanic islands or Antarctica. PA: Palaearctic, NA:
Nearctic, NT: Neotropical, AT: Afrotropical, OL: Oriental, AU: Australasian, PAC: Pacific Oceanc Islands, ANT: Antarctic
Hydrobiologia (2008) 595:219–224 223
123

highly modified member of the family Monodellidae
adapted to an exceptional habitat (hot springs; see
Wagner, 1994: 317).
Acknowledgement This is a contribution to Spanish MEC
project CGL2005-02217/ BOS.
References
Boutin, C. & P. Cals, 1985. Importance en biogeography
e
´
volutive de la de
´
couverte d’un Crustace
´
phre
´
atobie,
Monodella atlantomaroccana n. sp. (Thermosbaenacea)
dans la plaine alluviale de Marrakech (Maroc atlantique).
Comptes Rendus de l’Acade
´
mie des Sciences, Paris 300
(III, 7): 267–270.
Cals, P. & C. Boutin, 1985. De
´
couverte au Cambodge, do-
maine ancien de la Te
´
thys orientale, d’un nouveau
<<fossile vivant>> Theosbaena cambodjiana n.g., n.sp.
(Crustacea, Thermosbaenacea). Comptes rendus de
l’Acade
´
mie des sciences, (3) 300(8): 337–340.
Fryer, G., 1964. Studies on the functional morphology and
feeding mechanism of Monodella argentarii Stella
(Crustacea: Thermosbaenacea). Transactions of the Royal
Society of Edinburgh 66: 49–90.
Gordon, I., 1957. On Spelaeogriphus, a new cavernicolous
crustacean from South Africa. Bulletin of the British
Museum (Natural History), (Zoology) 5: 31–47.
Gordon, I., 1960. On a Stygiomysis from the West Indies, with
a note on Spelaeogriphus (Crustacea, Peracarida). Bulletin
of the British Museum (Natural History), (Zoology) 6:
285–326.
Pires, A. M. S., 1987. Potiicoara brasiliensis: a new genus and
species of Spelaeogriphacea (Crustacea: Peracarida) from
Brazil, with a phylogenetic analysis of the Peracarida.
Journal of Natural History 21: 225–238.
Poore, G. C. B. & W. F. Humphreys, 1998. First record of
Spelaeogriphacea from Australasia: a new genus and
species from an aquifer in the arid Pilbara of Western
Australia. Crustaceana 71: 721–742.
Poore, G. C. B. & W. F. Humphreys, 2003. Second species
of Mangkurtu (Spelaeogriphacea) from north-western Aus-
tralia. Records of the Western Australian Museum 22: 67–74.
Richter, S. & G. Scholtz, 2001. Phylogenetic analysis of the
Malacostraca (Crustacea). Journal of Zoological System-
atics and Evolutionary Research 39: 113–136.
Schram, F. R., 1974. Paleozoic Peracarida of North America.
Fieldiana Geology 33: 95–124.
Shen, Y.-B., F. R. Schram & R. S. Taylor, 1999. Liaoningo-
griphus quadripartitus (Malacostraca: Spelaeogriphacea)
from the Jehol Biota and notes on its paleoecology.
Palaeoworld 11: 175–187 (in Chinese).
Shen, Y. -B., R. S. Taylor & F. R. Schram, 1998. A new
spelaeogriphacean (Crustacea: Peracarida) from the Upper
Jurassic of China. Contributions to Zoology 68: 19–35.
Wagner, H. P., 1994. A monographic review of the Thermos-
baenacea (Crustacea: Peracarida). Zoologische Ver-
handelingen 291: 1–338.
224 Hydrobiologia (2008) 595:219–224
123