Албертс Б., Брей Д. и др. Молекулярная биология клетки. Том 1
Подождите немного. Документ загружается.


391
лоты внутрь клетки за собой. Чем выше градиент Na
+
, тем больше скорость всасывания растворенных молекул. Наоборот, если концентрация Na
+
во внеклеточной жидкости заметно уменьшается, - транспорт растворенных молекул останавливается. Гипотетическая (и довольно упрощенная)
схема функционирования подобной системы симпорта изображена на рис. 6-51.
У бактерий и растений большинство систем активного транспорта, приводящихся в действие ионными градиентами, используют в
качестве котранспортируемого иона Н
+
, а не Na
+
. В частности, активный транспорт большей части Сахаров и аминокислот в бактериальные клетки
обусловлен градиентом Н
+
через плазматическую мембрану. Наиболее хорошо изученный пример такого рода - переносчик лактозы (пермеаза).
Этот трансмембранный белок, состоящий из одной полипептидной цепи (длиной около 400 аминокислотных остатков), по-видимому, пересекает
липидный бислой по крайней мере девять раз. Он осуществляет Н
+
-зависимый симпорт: с каждой транспортируемой в клетку молекулой лактозы
переносится один протон.
6.4.10. Антипорты в плазматической мембране регулируют внутриклеточное значение рН [27]
Почти все клетки позвоночных имеют в составе плазматической мембраны (Na
+
+ Н
+
)-переносчик-обменник. Он играет ключевую роль
в поддержании внутриклеточного значения рН (pH
i
) обычно около 7,1 — 7,2. Этот переносчик обеспечивает сопряжение выброса ионов Н
+
с
притоком ионов Na
+
и, таким образом, удаляет избыток ионов Н
+
, образующийся в результате клеточных реакций окисления. Работа (Na
+
+ Н
+
)-
обменника регулируется значением рН: например, когда рН
i
в мышечных клетках цыпленка выше уровня 7,7 - обменник становится неактивным;
как только значение Ph
i
падает - активность обменника увеличивается, достигая половины своей максимальной активности при рН
i
7,4. Такая
регуляция обусловлена связыванием Н
+
с регуляторным участком обменника, находящимся на цитоплазматической стороне мембраны. Важность
(Na
+
+ К
+
)-обменника в поддержании уровня рН
i
была продемонстрирована экспериментами с мутантами фибробластов, не содержащими такого
обменника: они очень быстро погибали при помещении их в достаточно кислые условия, хотя нормальные фибробласты в этих условиях
жизнеспособны. В поддержании уровня рН
i
у многих ядерных клеток, по-видимому, играет важную роль и (С1
-
+ НСО
3
)-обменник, сходный с
белком полосы 3 из мембран эритроцитов (см. разд. 6.2.6). Подобно (Na
+
+ Н
+
)-обменнику, работа (Сl
-
+ НСО
3
-
)-обменника регулируется значением
рН
i
, но противоположным образом. Его активность возрастает при повышении рН
i
(т. е. когда цитозоль становится слишком щелочным),
увеличивая скорость выведения НСОз
-
из клетки в обмен на С1
-
и, таким образом, понижая рН
i
.
Существуют данные, свидетельствующие о том, что (Na
+
+ Н
+
)-обменник может участвовать не только в поддержании рН
i
, но и в
преобразовании внеклеточных сигналов во внутриклеточные. Например, большинство белковых факторов роста в процессе стимуляции клеточной
пролиферации активируют такого рода системы антипорта, увеличивая рН
i
от 7,1 или 7,2 до примерно 7,3. По крайней мере в некоторых случаях
они делают это косвенным образом, активируя cпецифическую протеин-киназу (протеинкиназа С - см. разд. 12.3.10), которая в свою очередь
фосфорилирует обменник. Это приводит к увеличению сродства регуляторного участка, связывающего Н
+
, и, следовательно, обменник остается
активным и при больших рН. Мутантные клетки, лишенные (Na
+
+ + Н
+
)-обменника, или клетки, обработанные препаратом амилоридом,
392
ингибирующим его, не способны к ответу на действие факторов роста. Все эти факты наводят на мысль, что активация обменника с последующим
увеличением рН
i
играет важную роль в инициации клеточной пролиферации. По-видимому, сходным образом увеличение рН
i
при оплодотворении
яиц морского ежа, вызванное активацией (Na
+
+ Н
+
)-обменника, стимулирует синтез ДНК и белка. Остается неясным, какие внутриклеточные белки
ответственны за увеличение рН
i
в ответ на эту активацию.
6.4.11. В основе межклеточного транспорта растворенных веществ лежит асимметричное распределение белков-переносчиков в
клетках эпителия [28]
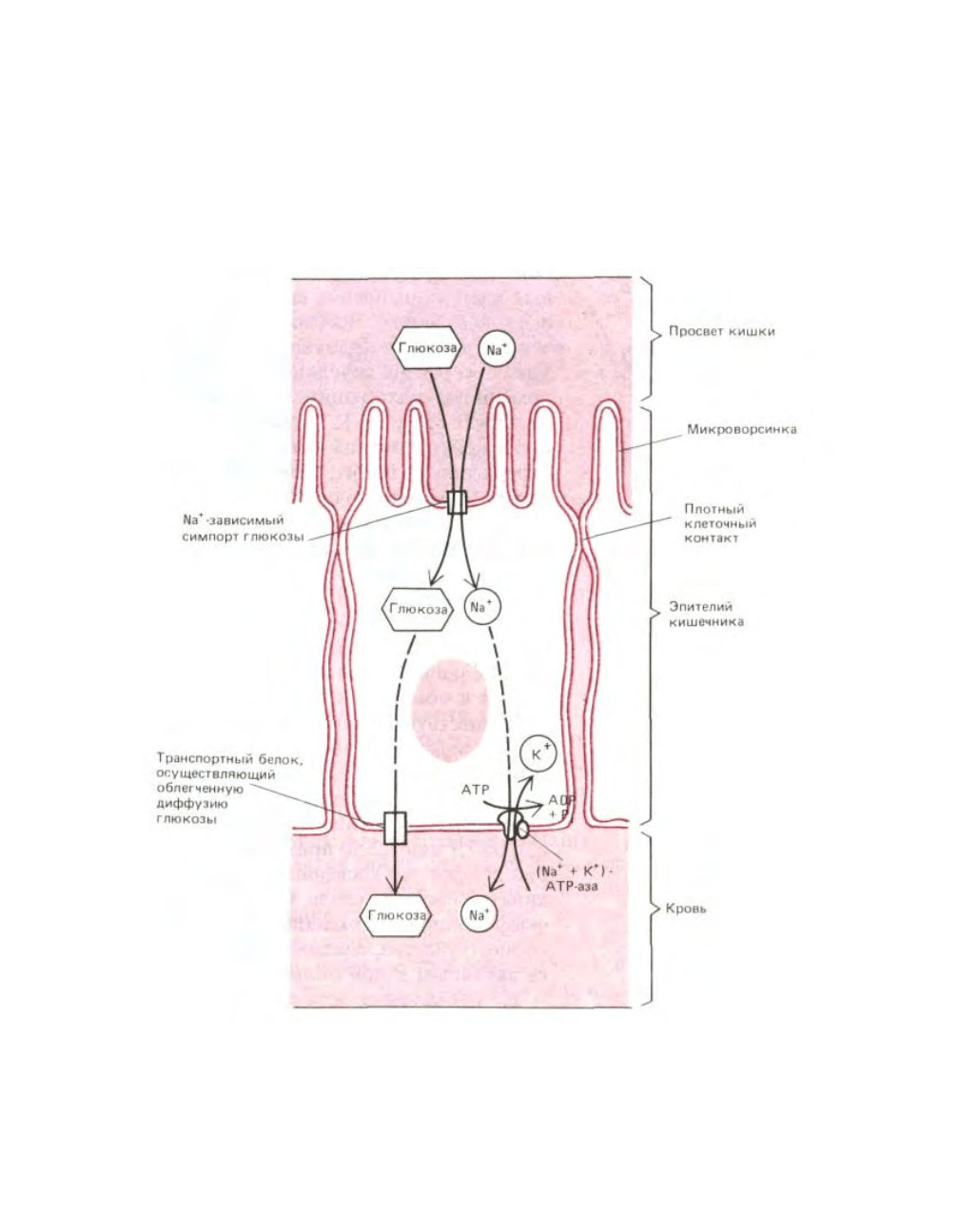
В плазматической мембране некоторых эпителиальных клеток, участвующих в поглощении пищи в кишечнике, белки-переносчики
распределены асимметрично, способствуя, таким образом, сквозному транспорту поглощенных растворенных веществ через клетку. Как показано
на рис. 6-52, белки, локализованные в плазматической мембране на апикальной (всасывающей) поверхности эпителиальной клетки, осуществляют
Na
+
-зависимый симпорт, перенося питательные вещества внутрь клетки. В то же время Na
+
-независимые белки в базальной и латеральной
мембранах позволяют питательным веществам выходить наружу по градиенту их концентраций. Градиент Na
+
на плазматической мембране таких
клеток поддерживается (Na
+
+ К
+
)-АТРазой, находящийся в базолатеральной области. По-видимому, сходные механизмы используются
эпителиальными клетками кишечника и почек для перекачивания молекул воды из одного внеклеточного пространства в другое.
Рис. 6-52. Асимметричное распределение транспортных белков в плазматической мембране клетки эпителия кишечника приводит к сквозному
транспорту глюкозы из полости кишки сквозь клетку во внеклеточную жидкость (откуда она поступает в кровь). Глюкоза проникает в клетку через
апикальную мембрану посредством Na
+
-зависимого симпорта глюкозы и выходит из клетки (по градиенту своей концентрации) путем облегченной
диффузии, опосредуемой другим белком-переносчиком глюкозы, локализованном в базальном и латеральном доменах. Градиент Na
+
, который
приводит в движение симпорт глюкозы, поддерживается (Na
+
+ К
+
)-АТРазой, находящейся в плазматической мембране базолатерального домена;
благодаря этому ферменту внутриклеточная концентрация Na
+
сохраняется на низком уровне. Соседние клетки соприкасаются между собой,
образуя непроницаемые контакты (называемые плотными контактами), обладающие двоякими функциями в представленном здесь процессе
транспорта. Эти контакты препятствуют проникновению растворов в щели между клетками эпителия и способствуют возникновению градиента
глюкозы через слой эпителиальных клеток. Плотные контакты также служат барьером внутри плазматической мембраны, препятствующим
диффузии молекул по поверхности мембраны. Благодаря им различные белки-переносчики удерживаются в соответствующих мембранных доменах
(см. рис. 6-36).
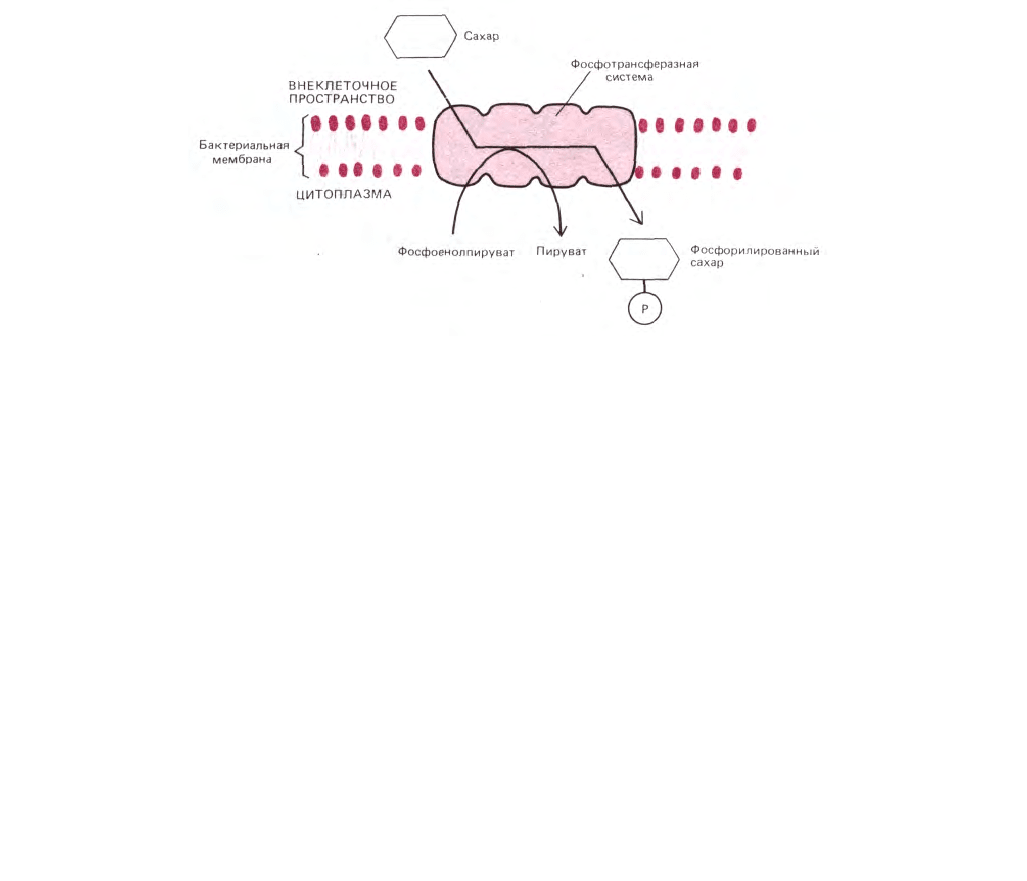
393
Рис. 6-53. Активный транспорт Сахаров внутрь бактериальных клеток за счет направленного переноса групп. Специальная «фосфотрансферазная
система белков» в бактериальной мембране фосфорилирует сахар сразу после переноса его через мембрану. Донором фосфата служит
фосфоенолпируват, а не АТР.
Во многих эпителиальных клетках площадь плазматической мембраны намного увеличена за счет существования тысяч микроворсинок,
выступающих с апикальной поверхности в виде тонких пальцевидных образований (рис. 6-52). Такие микроворсинки могут увеличивать общую
площадь всасывающей поверхности в 25 раз, тем самым значительно повышая транспортные возможности клетки. Апикальная поверхность
эпителиальной клетки кишечника является также местом, где локализованы иммобилизованные гидролитические ферменты, участвующие в
конечных стадиях переваривания пищи. Увеличение площади поверхности эпителия за счет микроворсинок в значительной степени способствует
перевариванию и всасыванию пищи.
6.4.12. Активный транспорт в бактериях может идти путем векторного переноса групп [29]
Как мы теперь знаем, активный транспорт в клетках может приводиться в действие светом (например, в бактериородопсине), гидролизом
АТР или же ионными градиентами. Четвертый путь, используемый некоторыми бактериями, состоит в том, чтобы молекулу, вошедшую в клетку
посредством пассивного транспорта, «поймать в капкан» с помощью химической модификации, которая не дает возможности молекуле выйти
обратно тем же путем. Например, сахара после переноса через плазматическую мембрану некоторых бактерий фосфорилируются. В результате
такой модификации они становятся заряженными, не могут выйти обратно и поэтому накапливаются в клетке. Более того, вследствие
фосфорилирования транспортируемых Сахаров концентрация их нефосфорилированных аналогов внутри клетки остается очень низкой, так что
градиент концентрации сахара продолжает «вталкивать» эти молекулы в клетку. Поскольку фосфатные группы переносятся на молекулы
растворенных веществ после их транспорта, этот тип активного транспорта называют векторным или направленным переносом групп. В
большинстве хорошо изученных примеров механизм фосфорилирования достаточно сложен и тщательно регулируется. В нем принимают участие
по крайней мере четыре отдельных мембранных белка, а в качестве донора высокоэнергетического фосфата используется фосфоенолпируват, а не
АТР (рис. 6-53).
6.4.13. Гактерии с двойными мембранами обладают транспортными системами, которые зависят от водорастворимых субстрат-
связывающих белков [30]
Как уже упоминалось выше, плазматические мембраны всех бактерий содержат белки-переносчики, использующие градиент ионов Н
+
для
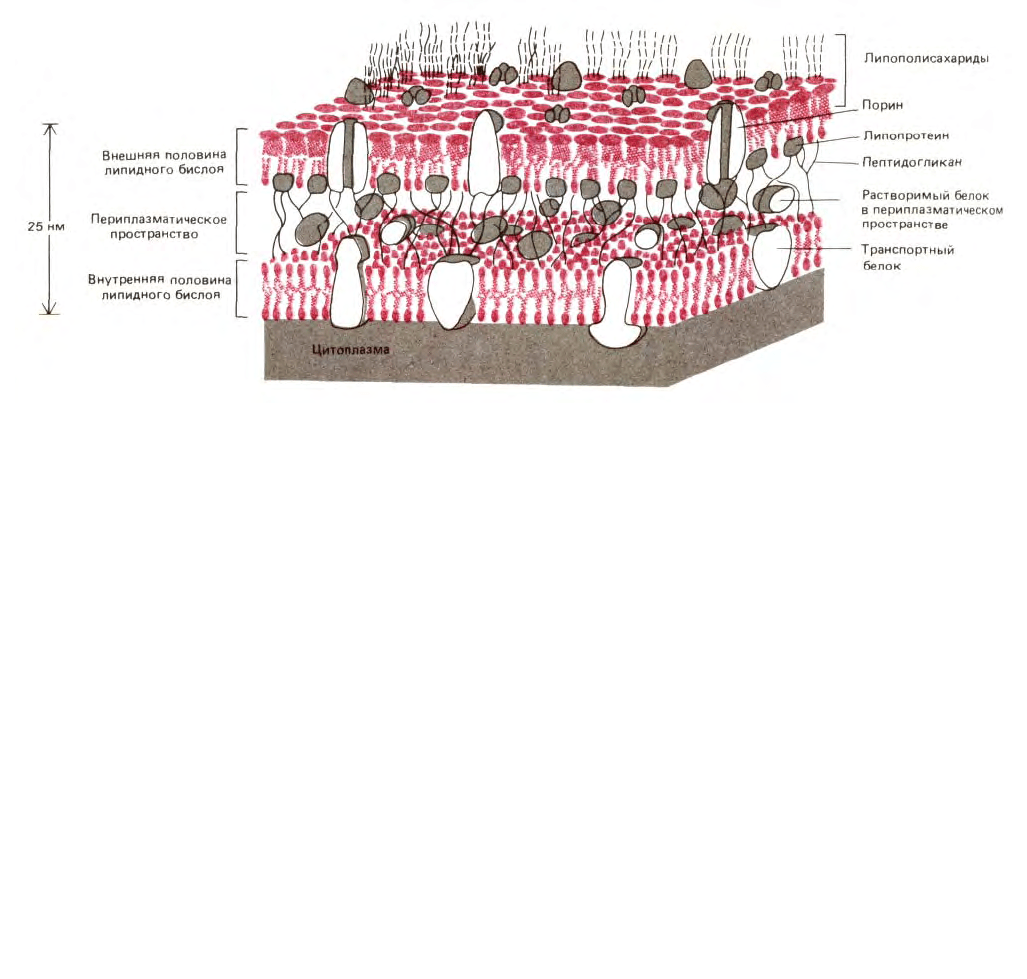
394
Рис. 6-54. Схематическое изображение небольшого участка двойной мембраны бактерий Е. соli. Внутренняя мембрана -это клеточная
плазматическая мембрана. Между внутренней и внешней липидными бислойными мембранами находится высокопористый, упругий
пептидогликан, состоящий из белков и полисахаридов, входящих в клеточные стенки бактерий. Он присоединен к липопротеиновым молекулам
внешней мембраны и заполняет периплазматическое пространство. В нем содержатся различные растворимые белки. Темные нити на
поверхности внешней мембраны представляют собой полисахаридные цепи специальных липополисахаридных молекул, образующих внешний
монослой верхней мембраны. Для простоты показаны лишь несколько цепей. Бактерии с двойной мембраной называются грамотрицателъными,
поскольку они не окрашиваются темно-синей краской по Граму. Бактерии с одной мембраной (но с толстыми клеточными стенками), например,
стафиллококки или стрептококки, окрашиваются по Граму и называются грамположительными. Их единственная мембрана аналогична внутренней
мембране грамотрицательных бактерий.
перекачки различных питательных веществ внутрь клетки. Однако многие бактерии, включая Е. coli, имеют еще и окружающую их внешнюю
мембрану, через которую растворенные вещества с молекулярной массой до 600 дальтон могут относительно свободно проникать через различные
каналообразующие белки (известные под общим названием порины) (рис. 6-54). У этих бактерий для переноса некоторых Сахаров, аминокислот и
мелких пептидов через внутреннюю (плазматическую) мембрану существует двухкомпонентная транспортная система, использующая
водорастворимые белки, локализованные в периплазматическом пространстве между двумя мембранами. Такие периплазматические субстрат-
связывающие белки присоединяют к себе специфические молекулы, которые нужно перенести через мембрану. При этом конформация белков
меняется, что позволяет им связываться с другим компонентом транспортной системы - трансмембранным белком-переносчиком, находящимся во
внутренней мембране (рис. 6-55). По-видимому, субстрат-связывающие белки передают присоединенное к ним растворенное вещество
специальному белку-переносчику, который затем использует энергию гидролиза АТР для переноса его через внутреннюю мембрану. Те же самые
периплазматические субстрат-связывающие белки служат и в качестве рецепторов в хемотаксисе -адаптивном процессе, позволяющем бактериям
перемещаться в направлении более высокой концентрации специфического питательного вещества.
Теперь же мы вновь обратимся к каналообразующим белкам.
6.4.14. Белковые каналы образуют в плазматической мембране поры [31]
В отличие от белков-переносчиков белковые каналы (или каналообразующие белки) формируют в мембранах поры, заполненные
водой. При этом каналообразующие белки внешних мембран бактерий (а также митохондрий и хлоропластов) образуют большие, относительно
неспецифичные поры, а в плазматических мембранах животных и растительных клеток эти поры малы по размеру и высоко специфичны. Почти все
белковые каналы служат для специфического транспорта ионов и обсуждаются здесь под названием ионных каналов. Ионные каналы
обеспечивают перенос приблизительно 10
6
ионов в секунду, что более чем в 100 раз больше скорости транспорта, осуществляемого любым из
известных белков-переносчиков. Ионные каналы никогда не работают совместно
395
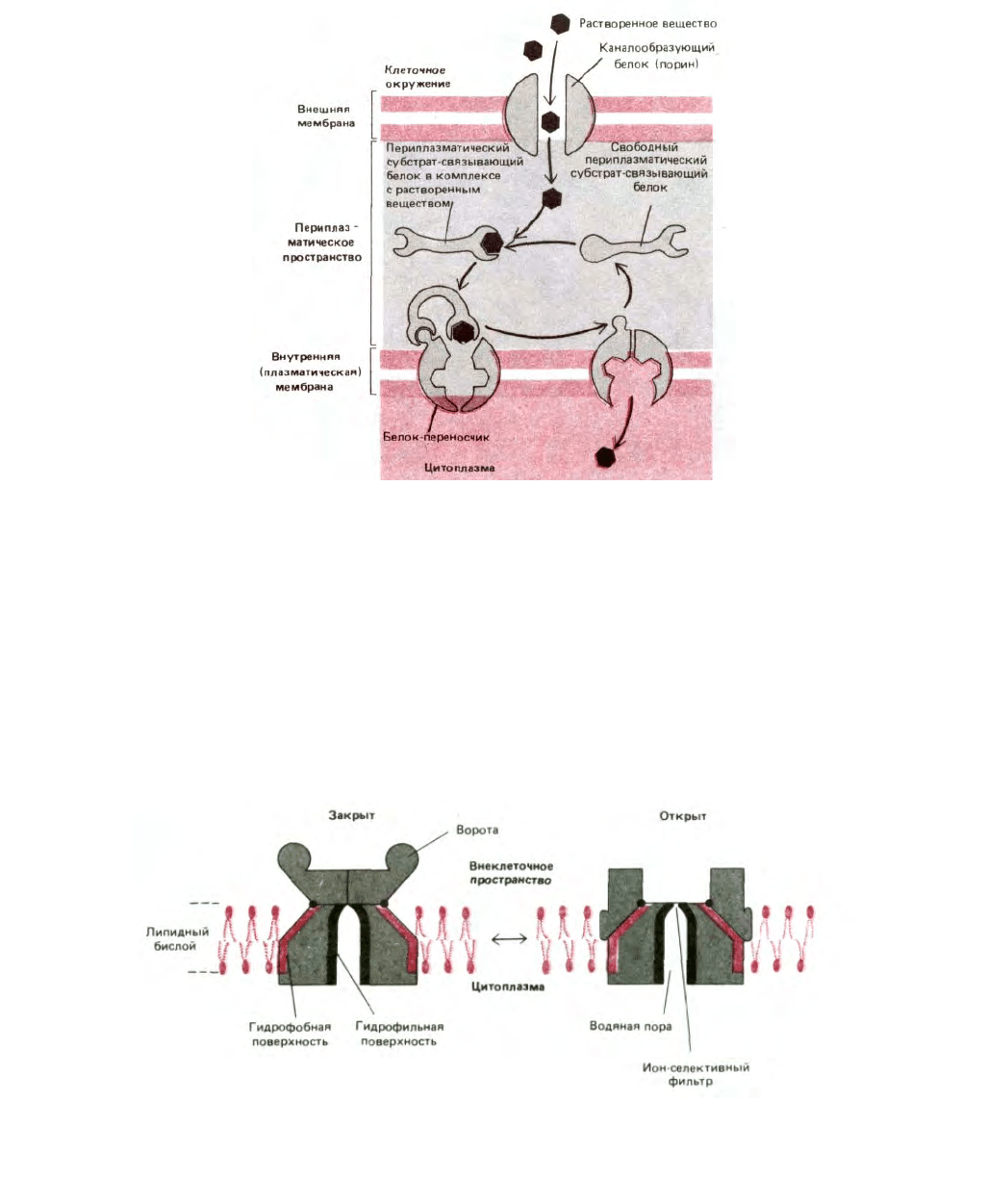
Рис. 6-55. Транспортная система, зависящая от периплазматических субстрат-связывающих белков в бактериях с двойной мембраной.
Растворенные вещества диффундируют через каналообразующие белки (порины), находящиеся во внешней мембране, и связываются с
периплазматическими субстрат-связывающими белками. При этом белки испытывают конформационные изменения, приобретая способность
связываться с белками-переносчиками плазматической мембраны, которые затем перехватывают субстрат и активно транспортируют его через
бислой. Эта стадия опосредуется гидролизом АТР. Пептидогликаны для простоты не показаны. Их пористая структура позволяет субстрат-
связывающим белкам и водорастворимым веществам двигаться путем простой диффузии.
с источником энергии; осуществляемый ими транспорт всегда пассивный («с горки») и позволяет специфическим ионам, главным образом Na
+
, К
+
,
Са
2+
или С1
-
, диффундировать по их электрохимическим градиентам через липидный бислой.
Белковые каналы плазматической мембраны обладают ионной селективностью, т. е. позволяют диффундировать через них только ионам
определенного вида. По-видимому, поры должны быть достаточно узкими, чтобы ионы находились в тесном контакте с их стенками и чтобы
проходить могли только те из них, которые имеют подходящий размер и заряд. Скорее всего на этом пути ионам приходится терять большинство
или даже все ассоциированные с ними молекулы воды. Эти два обстоятельства накладывают ограничение на скорость диффузии через канал и
делают его селективным фильтром, допускающим прохождение только ионов определенного типа. Таким образом, при увеличении концентрации
ионов их поток через канал возрастает пропорционально, но лишь до определенного предела.
Другой особенностью, отличающей ионные каналы от простых пор, заполненных водой, является то, что они открыты не все время. Как
показано на рис. 6-56, каналы имеют «ворота», которые открываются на
Рис. 6-56. Схематическое изображение воротного ионного канала в закрытой и открытой конформациях. Трансмембранный белок, показанный в
разрезе, образует в липидном бислое заполненную водой сквозную пору при открытых воротах. В состав стенок поры входят, видимо,
гидрофильные аминокислотные остатки. Гидрофобные остатки взаимодействуют с липидным бислоем. Ионная селективность канала определяется
самым узким его местом. Временное открытие ворот вызывается специфическим возмущением мембраны, различным для разных каналов
(обсуждается в тексте). Положение ворот и ионного селективного фильтра для большинства каналов неизвестно.

396
короткое время, а затем закрываются. В большинстве случаев ворота открываются в ответ на специфические возмущения мембраны. Среди них
наиболее известны в настоящее время изменение мембранного потенциала (потенциал-зависимые воротные каналы), механическая стимуляция
(механически открываемые каналы - см. разд. 19.6.3) или связывание сигнальных молекул (лиганд-зависимые воротные каналы). Сигнальными
лигандами могут быть либо внеклеточные посредники, называемые нейротрансмиттерами или нейромедиаторами (нейротрансмиттер-
зависимые каналы), либо внутриклеточные посредники, например ионы (ион-зависимые каналы - см. разд. 21.1.1.), нуклеотиды (нуклеотид-
зависимые каналы - см. разд. 3.3.5) или GTP-связывающие регуляторные белки (G-белок-регулируемые каналы - см. разд. 12.3.12).
На сегодняшний день известно уже около 50 видов ионных каналов. И в настоящее время продолжают открывать все новые их виды.
Ионные каналы ответственны за электрическую возбудимость нервных и мышечных клеток; они осуществляют большинство форм передачи
электрических сигналов в нервной системе. Отдельная нервная клетка обычно содержит более пяти видов ионных каналов. Однако такие каналы
присущи не только электрически возбудимым клеткам. Они есть у всех животных клеток и обнаруживаются в некоторых клетках растений и
микроорганизмов. Именно эти каналы, например, ответственны за закрывание листьев в ответ на раздражение у мимозы или за изменение
направления движения на обратное у одноклеточного Paramecium.
Видимо, наиболее распространенными ионными каналами являются те, которые проницаемы главным образом для К
+
. Они обнаружены
в плазматических мембранах почти всех животных клеток. Поскольку для их открывания скорее всего не требуется специфических мембранных
возмущений, их называют иногда калиевыми проточными каналами. Эти каналы играют ключевую роль в установлении мембранного потенциала -
разности электрического напряжения, наблюдающейся на двух сторонах всех типов мембран.
6-24
6-26
6.4.15. Мембранный потенциал зависит от К
+
-проточных каналов и градиента К
+
через мембрану [32]
Мембранный потенциал зависит от распределения ионов на обеих сторонах мембраны. Выше уже шла речь о том, что (Na
+
+К
+
)-АТРаза
способствует установлению осмотического равновесия в клетке за счет поддержания низкой внутриклеточной концентрации Na
+
. Из-за низкой
концентрации натрия внутри клетки необходим избыток других катионов, чтобы сбалансировать заряд фиксированных клеточных анионов -
отрицательно заряженных органических молекул, находящихся внутри клетки. Эту роль выполняют главным образом ионы калия благодаря К
+
-
проточным каналам, которые обеспечивают свободный переход этих ионов через мембрану и позволяют им засасываться внутрь клетки за счет
отрицательного заряда фиксированных анионов (даже при отсутствии какой бы то ни было работы (Na
+
+ К
+
)-АТРазы. Таким образом,
устанавливается равновесие, при котором электрическая сила, «втягивающая» ионы калия внутрь клетки, уравновешивается стремлением К
+
вытекать из клетки по градиенту концентрации. Мембранный потенциал является выражением этой электрической энергии и его величина может
быть рассчитана из крутизны градиента концентрации К
+
, необходимой для уравновешивания электрических сил. Поясним на таком примере.
Предположим, что электрический градиент через плазматическую мембрану первоначально отсутствует (т. е. мембранный потенциал равен нулю),
но концентрация К
+
внутри клетки выше (для уравновешивания

397
УРАВНЕНИЕ НЕРНСТА И ПОТОК ИОНОВ
Поток любых ионов через белковый мембранный канал зависит от их электрохимического градиента. Этот градиент представляет
собой комбинацию двух компонентов: градиента напряжения и градиента концентрации ионов через мембрану. Когда эти две составляющие
уравновешивают друг друга, электрохимический градиент для данного иона равен нулю. Это значит, что суммарный поток ионов через канал
будет отсутствовать. Градиент напряжения (мембранный потенциал), при котором наблюдается такое равновесие, называется потенциалом
равновесия. Он может быть вычислен из уравнения, которое будет выведено ниже и называется уравнением Нернста.
Уравнение Нернста
V = RT/ zF lnC
0
/C
i
,
где V — потенциал равновесия в вольтах (потенциал внутри клетки минус потенциал
снаружи),
C
0
и С
i
— внешняя и внутренняя концентрация ионов,
соответственно,
R — универсальная газовая постоянная (2 кал х моль
-1
х °К)
Т — абсолютная температура (°К),
F — постоянная Фарадея (2,3 х 10
4
кал х В
-1
х моль
-1
) ,
z — заряд иона.
Уравнение Нерста выводится следующим образом: молекулы в растворе всегда движутся из области с высокой концентрацией в
область с низкой. Следовательно, движение по градиенту концентрации сопровождается понижением свободной энергии (∆G < 0), тогда как
движение против градиента концентрации ведет к возрастанию свободной энергии (∆G > 0). (Понятие свободной энергии вводится и
обсуждается на схеме 2-7.). Изменение свободной энергии на моль растворенного вещества, перенесенного через мембрану (∆G
conc
) равно —
RT lnC
0
/C
i
. Если растворенным веществом являются ионы, то их перемещение внутрь клетки через мембрану (со скачком напряжения на ней,
равным V) будет вызывать дополнительное изменение свободной энергии (на моль перемещенного вещества) ∆G = -zFV. При условии
равновесия ∆G
conc
+ ∆G
volt
= 0 распределение ионов на двух сторонах Мембраны будет равновесным. Таким образом,
zF - RT lnC
0
/C
i
= 0,
и, следовательно,
V = RT/zF In C
0
/C
i
= 2,3 RT/zF Ig C
o
/C
i
.
Для одновалентных ионов
2,3 RT/F - 58 мВ при 20°С и 61,5 мВ при 37°С.
Поэтому, когда мембранный потенциал имеет значение 61,5 lg( [К
+
]
0
/ [К
+
],) милливольт (-89 мВ, когда [К
+
]
о
= 5 мМ и [К
+
] = 140
мВ), он является потенциалом равновесия для К
+
(V
k
), при котором отсутствует суммарный ток К через мембрану. Аналогично, когда
мембранный потенциал имеет значение 61,5 Ig ( [Na
+
]
o
/ [Na
+
]
i
) — равновесный потенциал для Na
+
(V
Na
), отсутствует ток Na
+
. Для типичной
клетки V
k
колеблется от — 70 мВ до — 100 мВ, a V
Na
от +50 мВ до +65 мВ.
Для какого-либо отдельного мембранного потенциала
V
M
, суммарная сила, стремящаяся вывести отдельный тип ионов из клетки, пропорциональна разнице между V
M
и равновесным
потенциалом для этих ионов. Например, для K
+
: V
M
— V
k
и для Na
+
: V
M
— V
Nа.
Реальный ток, осуществляемый каждым видом ионов зависит
не только от движущей силы, но также и от степени проницаемости мембраны для данного иона через каналы, которая есть функция
проводимости каналов для данного иона. Если проводимость набора каналов для К
+
и Na
+
соответственно g
К
и g
Na
, из закона Ома следует, что
токи К
+
и Na
+
соответственно будут равны g
К
( V
м
- V
к
) и g
Na
(V
M
- V
Na
) (пpoводимость есть величина, обратная сопротивлению, а единица
измерения, обратная ому, есть сименс, S). Большинство индивидуальных ионных каналов имеет проводимость в пределах 1 — 150 х 10
-12
S
или 1 — 150 pS). При мембранном потенциале покоя в большинстве клеток К -каналы являются основным типом открытых каналов. Поэтому
g
k
доминирует в общей проводимости мембраны. В процессе возникновения потенциала действия в нервных или мышечных клетках
открывается на короткое время множество потенциал-зависимых Nа
+
-каналов, в результате проводимость g
Na
становится доминирующей в
общей проводимости мембраны.
Схема 6-2. Вывод уравнения Нернста.
398
заряда фиксированных анионов), чем снаружи. В этом случае ионы калия будут стремиться выйти из клетки через проточные К
+
-каналы по
направлению градиента их концентрации. Если ионы выйдут из клетки, то внутри останется отрицательный заряд и, таким образом, возникнет
электрическое поле, другими словами, мембранный потенциал, стремящийся вернуть ионы К
+
в клетку. Вытекание ионов калия прекратится, как
только образовавшийся мембранный потенциал достигнет значения, при котором электрическая движущая сила, действующая на ионы калия, точно
уравновесит действие градиента концентрации К
+
, т.е. когда электрохимический градиент ионов калия окажется равным нулю. Таким же образом
создается одновременно и равновесие для ионов Сl
-
, но, поскольку их заряд отрицательный, они удерживаются вне клетки. Равновесные условия,
при которых отсутствует электрический ток через мембрану, определяют клеточный мембранный потенциал покоя. Существует простая, но
очень важная формула, количественно выражающая условия равновесия - уравнение Нернста. Как показано на схеме 6-2, она позволяет
рассчитать мембранный потенциал покоя, если известно соотношение внутренней и внешней концентраций ионов.
Для того чтобы установился мембранный потенциал, достаточно перенести через мембрану совсем небольшое количество ионов. Таким
образом, мембранный потенциал можно представить себе как перемещение зарядов, оставляющее концентрации ионов практически неизменными.
В результате происходит лишь небольшое перераспределение числа положительно и отрицательно заряженных ионов между сторонами мембраны
(рис. 6-57). Более того, это перемещение зарядов происходит очень быстро, за несколько миллисекунд или даже быстрее.
Рассмотрим, что случится, если инактивировать (Na
+
+ К
+
)-АТРазу. Прежде всего произойдет небольшое быстрое падение мембранного
потенциала, поскольку (Na
+
+ К
+
)-насос является электрогенным и в активном состоянии вносит свой вклад в мембранный потенциал (см. разд.
6.4.6). Однако выключение этого насоса не приводит к исчезновению главного компонента потенциала покоя, основанного на механизме
уравновешивания ионами калия (как описано выше). Он существует до тех пор, пока концентрация Na
+
внутри клетки остается низкой, т.е. многие
минуты. Но поскольку плазматическая мембрана хоть и плохо, все же проницаема для ионов Na
+
, то Na
+
будет медленно входить внутрь клетки по
своему электрохимическому градиенту. Приток натрия уменьшает мембранный потенциал и, таким образом, вызывает дополнительный отток
ионов К
+
из клетки. В это время нарушается осмотическое равновесие (см. разд. 6.4.6), но, если клетка не лопнула, со временем установится новое
состояние равновесия между ионами Na
+
, К
+
и С1
-
. При этом мембранный потенциал будет намного ниже, чем в нормальной клетке с активным
(Na
+
+ К
+
)-насосом.
Точное равенство зарядов на обеих сторонах
мембраны; мембранный потенциал = 0
Несколько положительно заряженных ионов (цветные) пересекли мембрану справа налево,
оставив с другой стороны отрицательно заряженные противоионы (цветные); при этом
мембранный потенциал отличен от нуля
Рис. 6-57. Небольшой поток ионов несет достаточный заряд для создания большого изменения мембранного потенциала. Ионы, определяющие
мембранный потенциал, располагаются вблизи мембраны, удерживаясь за счет взаимодействия с противоионами другой стороны мембраны. Для
типичной клетки 1 микрокулон заряда (6 х 10
12
одновалентных иона) на 1 см
2
мембраны, перенесенный с одной стороны на другую, изменит
мембранный потенциал примерно на I В. Это значит, что для сферической клетки диаметром 10 мкм вытекание из клетки лишь 1/100000 доли
ионов К
+
будет изменять потенциал на 100 мВ.
399
Разность потенциалов на сторонах плазматической мембраны клетки, находящейся в покое, варьирует в зависимости от организма или
типа клеток от — 20 мВ до — 200 мВ. Хотя градиент К
+
всегда вносит наибольший вклад в этот потенциал, значительным эффектом обладают
также и градиенты других ионов (плюс неравновесные эффекты ионных насосов). Чем более проницаема мембрана для данного иона, тем в
большей степени мембранный потенциал зависит от равновесных условий для этого иона. Следовательно, практически при любом изменении
проницаемости мембраны для ионов происходит изменение и мембранного потенциала. Это ключевой принцип, связывающий электрическую
возбудимость клеток с активностью ионных каналов.
6-25
6.4.16. Потенциал-зависимые воротные ионные каналы ответственны за электрическую возбудимость нервных и мышечных
клеток [33]
В плазматических мембранах электрически возбудимых клеток (главным образом нервных и мышечных) содержится множество
потенциал-зависимых воротных ионных каналов, ответственных за генерацию потенциалов действия - быстрых, скоротечных
самораспространяющихся электрических возбуждений мембраны. Этот процесс начинается при деполяризации мембраны - смещении мембранного
потенциала к менее отрицательному значению. Стимул, который вызывает моментальную частичную деполяризацию, сразу же открывает
потенциал-зависимые воротные Na
+
-каналы, что позволяет небольшому количеству Na
+
войти в клетку. Приток положительных зарядов в свою
очередь деполяризует мембрану еще больше, приводя к открыванию других Na
+
-каналов, пропускающих дополнительное количество ионов натрия
и, таким образом, дальнейшую деполяризацию. Этот процесс продолжается до тех пор, пока потенциал локального участка мембраны не изменится
от своего значения покоя около — 70 мВ до равновесного потенциала Na
+
, равного примерно + 50 мВ (см. схему 6-2). При этом значении, когда
суммарный электрохимический потенциал Na
+
равен нулю, клетки пришли бы в новое состояние равновесия (или покоя), в котором все натриевые
каналы перманентно открыты, если бы открытая конформация канала была стабильной. Однако клетки защищены от такого непрерывного
электрического спазма, поскольку Na
+
-каналы находятся под контролем автоматического инактивирующего механизма. Они быстро закрываются
после открытия, несмотря на деполяризацию мембраны. В таком инактивированном состоянии каналы не могут вновь открыться до тех пор, пока
не пройдет несколько миллисекунд после падения мембранного потенциала до первоначального отрицательного значения. На рис.
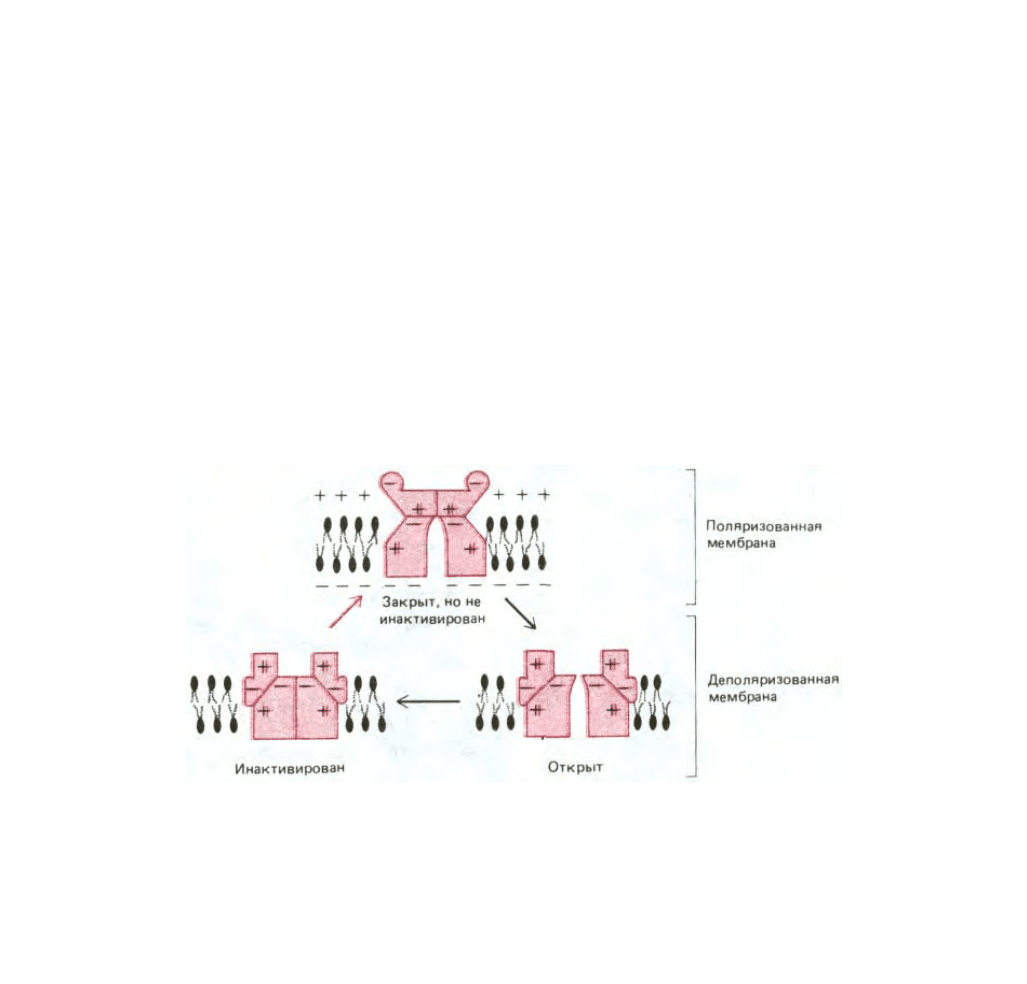
Рис. 6-58. Потенциал-зависимый Na
+
-канал может находиться в одном из трех по крайней мере состояний (конформаций). Внутренние силы,
представленные здесь в виде взаимодействия зарядов, находящихся на различных сторонах канала, стабилизируют каждое состояние и защищают
от влияния небольших возмущений. Однако взаимодействия с другими молекулами могут привести к переходу из одного состояния канала в
другое. Состояние с наименьшей энергией определяется мембранным потенциалом, так как различные конформаций имеют различное
распределение зарядов. В состоянии покоя (мембрана сильно поляризована) канал закрыт, но не инактивирован. Это наиболее стабильное
состояние с наименьшей свободной энергией. При деполяризации мембраны более низкой энергией будет обладать открытая конформация и,
следовательно, канал откроется. Но свободная энергия инактивированного состояния еще ниже и после некоторого случайного периода времени в
открытом состоянии канал переходит в инактивированное состояние. Таким образом, открытая конформация соответствует метастабильному
состоянию, существующему недолго. Черные стрелки показывают последовательность событий при деполяризации мембраны, а красная стрелка
обозначает возврат в первоначальное состояние с наименьшей свободной энергией после реполяризации мембраны.
400
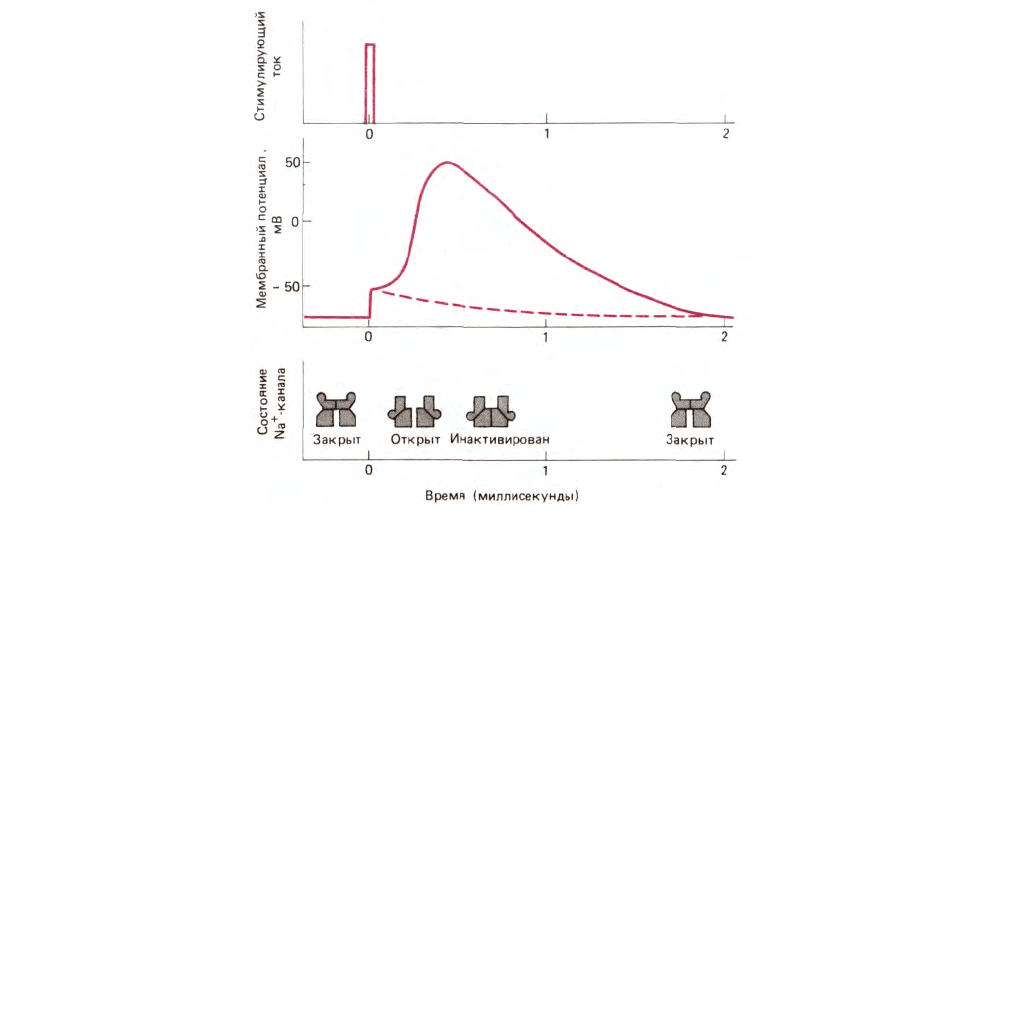
Рис. 6-59. Индукция потенциала действия коротким электрическим импульсом (показан на верхнем графике). Импульс частично деполяризует
мембрану (средний график). Сплошная линия на графике мембранного потенциала показывает возникновение потенциала действия при открывании
и последующей инактивации потенциал-зависимых Na
+
-каналов. Мембранный потенциал автоматически возвращается к своему первоначальному
значению — 70 мВ при закрытии Na
+
-каналов благодаря непрерывному вытеканию К
+
через К -каналы. Возникновение второго потенциала
действия невозможно до тех пор, пока Na
+
-каналы (их состояния показаны внизу) не вернутся в закрытое, но не инактивированное состояние (см.
рис. 6-58). До этого мембрана остается нечувствительной к раздражению. Прерывистая линия показывает релаксацию мембранного потенциала при
слабых воздействиях, не приводящих к открыванию каналов.
6-58 схематически изображены эти три различных состояния потенциал-зависимого воротного Na
+
-канала - закрытое, но не инактивированное,
открытое и инактивированное. Рис. 6-59 показывает, как этот канал работает при повышении и понижении потенциала действия.
Рассмотренный выше процесс образования потенциала действия относится лишь к небольшому участку плазматической мембраны.
Однако самоусиливающейся деполяризации этого участка достаточно для деполяризации соседних областей мембраны, которые вовлекаются при
этом в тот же цикл генерации потенциала действия. Таким образом, потенциал действия распространяется от первоначального участка
деполяризации по всей плазматической мембране. Более детальное рассмотрение функций и свойств потенциала действия приводится в гл. 19.
В нейронах и мышечных клетках имеется несколько тысяч потенциал-зависимых Na
+
-каналов. Ток, протекающий через мембрану, - это
сумма всех микротоков через каждый канал. Суммарный ток можно записать с помощью микроэлектрода (см. разд. 19.2.3). Однако существует
возможность зарегистрировать и токи, протекающие через индивидуальные каналы. Это делают с помощью специальной техники, позволяющей
выделить совсем небольшой участок мембраны, который содержит лишь несколько каналов, и записать затем протекающие через него токи. Такая
методика (patch-clamp) дает более детальную картину работы этих каналов.
6-27
6.4.17. Регистрация токов, проходящих через изолированный участок мембраны, показывает, что индивидуальные Na
+
-каналы
открываются по принципу «все или ничего» [34].
Разработка метода patch-clamp позволила существенно продвинуться в изучении ионных каналов. С помощью этого метода можно
анализировать транспорт через единичную молекулу белкового канала, находящегося на маленьком участке мембраны (рис. 6-60), записывать
сигналы