Васильев А.А. Теоретическая биология
Подождите немного. Документ загружается.

71
Таким образом, можно ожидать, что для «лучших» осмотиков комбинация (13) равна
0,5/120 г.
Отметим, что рациональное использование «хороших» осмотиков предполагает возмож-
ность обойтись весьма небольшими их количествами, т.е. использовать «хорошие» осмотики
весьма экономно, накапливая их только в зонах активно происходящих физиологических про-
цессов. И наоборот, использование «хороших» осмотиков не требуется в клеточных компар-
тментах, где процессы происходят не столь активно. Таким компартментом, прежде всего, яв-
ляется вакуоль как составляющая большую часть объема клетки и при этом относительно
инертная ее часть, в которой могут быть использованы «худшие» осмотики, в т.ч. неорганиче-
ские ионы и отходы жизнедеятельности в виде органических молекул или их низкомолекуляр-
ных производных. Описываемая возможность объясняет то обстоятельство, что абсолютные
количества веществ, предположительно используемых для осмотической адаптации и пере-
численных в таблице (в частности, наиболее часто обнаруживаемых –– пролина и глицинбе-
таина), оказываются невелики по сравнению с общей сухой массой тканей растения (хотя на
порядки превосходят таковое в сравнении с отсутствием засоления). И обратно, неэкономное
использование «хорошего» осмотика (синтез его в б0льших количествах, чем необходимо для
осуществления процессов в активно функционирующих компартментах клетки) можно рас-
сматривать как фактор несовершенства системы осмотической адаптации.
Естественно ожидать, что менее совершенная система осмотической адаптации будет ха-
рактерна для объектов и ситуаций, для которых долговременная и однозначно ориентирован-
ная на преодоление ограничений в связи с водным режимом стратегия не является насущно
необходимой (т.е. если достижение предельных возможностей осмотической адаптации не
является необходимым условием для нормального физиологического развития). В частности,
можно ожидать, что заведомо не самой эффективной с точки зрения предельных возможно-
стей механизма осмотической адаптации будет реакция на кратковременно действующий вод-
ный дефицит.
А именно, ожидаемая реакция при кратковременной осмотической адаптации состоит в
мобилизации наиболее доступного, но не самого подходящего для обеспечения физиологиче-
ских процессов <в долговременном аспекте> осмотика, который обеспечивает только текущий
водный баланс. Долговременная реакция состоит в замене исходно наиболее доступного осмо-
тика на более подходящий специально синтезируемый осмотик.
Как распространенный вариант кратковременной осмотической адаптации можно пред-
ставить адаптацию к изменению водного режима при обычной картине суточного изменения
условий. Осмотическая адаптация нужна в середине дня, когда дефицит паров воды в воздухе
достигает максимального значения.
В этом случае для адаптации целесообразно использование низкомолекулярных продук-
тов фотосинтеза, несмотря на то, что гексозы и тем более сахароза имеют плохие осмотиче-
ские свойства по всем показателям. Молекулярная масса этих соединений относительно вели-
ка. Их электростатическая конфигурация очень активна. А их синтез из-за больших затрат
энергии нежелателен не только с экономической, но и с экологической точки зрения, т.к. важ-
но, чтобы используемые соединения были менее привлекательны для травоядных.
Тем не менее, при использовании сахаров осмотическую адаптацию можно обеспечить
автоматически как побочный результат фотосинтеза почти без дополнительных затрат. Асси-
миляты в любом случае рационально накапливать в листе, чтобы было чем загрузить систему
флоэмного транспорта ночью. Поэтому днем, когда возникает потребность в осмотической
адаптации, а избыток ассимилятов в листе уже имеется, не держать этот избыток в форме
крахмала даже выгоднее –– не придется затрачивать энергию в цикле синтез/гидролиз крахма-
ла.
Суть изменений, происходящих при долговременной ОА можно представить как переход
от исходного состава высокомолекулярных и низкомолекулярных составляющих к конечному,
при котором с увеличением количества осмотиков в единице объема происходит пропорцио-
нальное уменьшение количества всех составляющих, обеспечивающих отдельные микроско-
пические этапы физиологических процессов. Общее количество вещества в единице объема
72
при заданной плотности ограничено (см. выше), поэтому должно происходить уменьшение
количества не только низкомолекулярных <вносящих наибольший вклад в осмотическое дав-
ление>, но высокомолекулярных составляющих.
Для описания такого изменения удобно ввести общий для этих составляющих масштаб-
ный коэффициент fm: в силу приближенной стехиометрии превращений и экономической це-
лесообразности (п.2.3–4) естественно ожидать, что примерно пропорционально fm будут из-
меняться максимальные и действующие скорости всех процессов.
Предельное значение масштабного коэффициента линейно уменьшается с увеличением
количества осмотиков. В силу экономической целесообразности следует ожидать, что наблю-
даемое значение fm будет близко к предельно возможному при заданном количестве осмоти-
ков. Учитывая линейную связь количества осмотиков с водным потенциалом, ожидаемую
связь значения fm с регистрируемым значением водного потенциала листа можно представить
в виде:
f
m
/f
m0
+ а |ψ
l
| = 1, (14)
где значение fm0 дает предельно возможное (максимальное) значение fm в отсутствии осмоти-
ческих ограничений, а коэффициент а = 1/πкрит зависит от типа осмотиков (их задерживаю-
щих свойств, выражаемых в общем случае соотношением (13)). Для «худшего» осмотика
больше значение а <т.е. при меньшем ψl происходит большее вытеснение физиологических
процессов>, а «лучший» осмотик отличается от «худших» тем, что вытесняет другие состав-
ляющие в наименьшей степени.
Таким образом, независимо от того, насколько совершенна используемая система осмо-
тической адаптации, любые используемые для осмотической адаптации вещества (осмотики)
вытесняют другие составляющие из полезного объема. Иными словами, рассматриваемую ги-
потетическую модель осмотической адаптации можно было бы назвать моделью инертного
вытеснения, имея в виду, что присутствие используемых при осмотической адаптации веществ
означает вытеснение других клеточных составляющих, непосредственно участвующих в фи-
зиологических процессах.
Формально выражение (14) действительно позволяет описать инертное вытеснение (то-
гда nкрит почти не зависит от типа осмотиков, т.е. при описании действия различных осмоти-
ков можно игнорировать зависимость nкрит({cj}) и использовать типовое n), но фактически
возможности применения этого выражения шире. Такое же по виду выражение опишет и ожи-
даемый результат адаптации при любом другом неспецифическом <как оказываемое на все
или многие процессы> деструктивном действии, лишь бы кооперативным был эффект этого
действия и механизм адаптации к нему.
Например, кооперативным деструктивным эффектом является действие некоторых неор-
ганических ионов на структуру макромолекул. Таково, в частности, действие, приводящее к
изменению пространственной структуры ферментов, и как следствие, потере (или заметному
уменьшению) проявляемой ими каталитической активности. Тогда будем наблюдать анало-
гичную критическую S-образную зависимость активности ферментов от водного потенциала,
линейно связанного с концентрацией ионов. Возможный кооперативный механизм адаптации
к такому деструктивному действию состоит в появлении частиц, образующих комплексы с
ионами. Другой вариант –– появление частиц, образующих комплексы с ферментами и предо-
храняющими их от связывания с ионами. И в том, и в другом случае будут выражены линей-
ные ограничения активности в связи с водным потенциалом.
Кроме того, выражение (14) опишет также кооперативное деструктивное действие в том
случае, если какой-либо механизм адаптации к нему не действует, например, не активирован,
как в случае, описанном на рисунке 6 для неадаптированных к водному дефициту растений.
Спад в критической области происходит почти линейно, а поэтому выражение вида (14) впол-
не подходит для его описания.
Поэтому выражаемое уравнением (14) предположение следует определить шире –– как
модель неспецифического кооперативного действия. При более широкой трактовке nкрит мо-
жет существенно отличаться от характерного, наблюдаемого только при инертном вытесне-
нии. А именно, значение nкрит должно быть больше характерного при инертном вытеснении,
73
т.к. по сути дела речь идет о выполнении более чем одного соотношения вида (14), которые
вместе эквивалентны одному, выражающему самое жесткое ограничение. Например, рассмот-
ренное выше кооперативное действие ионов на структуру макромолекул возможно наблюдать
через уменьшение скорости физиологических процессов только в том случае, когда ему соот-
ветствует большее nкрит по сравнению с характерным без такого действия –– в противном
случае процессы прекратятся при меньшем ψ из-за недостатка собственно водной среды.
Согласование с предшествующими этапами экономического описания
Описание ограничений в связи с осмотической адаптацией можно отнести к любой ткани
и органу растения, но наиболее актуальны такие ограничения по отношению к листу как ак-
тивно транспирирующему органу и по отношению к фотосинтезу как основному физиологиче-
скому процессу в листе, к тому же сопряженному с транспирацией. Поэтому утверждения об
осмотической адаптации наиболее естественно формулировать и проверять в связи с фотосин-
тезом.
Имеющиеся данные по влиянию на фотосинтез факторов, различным образом связанных
с водным режимом, значительно менее однозначны, чем уже рассмотренные данные по усть-
ичной регуляции фотосинтеза или данные по регуляции соотношения максимальных скоро-
стей световых и темновых этапов фотосинтеза. В связи с водным режимом наблюдается суще-
ственно более сложная картина в сравнении с представленной в пп.5–6 и приложении C, когда
при действии какого-либо одного фактора (при том, что общее число рассмотренных факторов
было весьма велико) качественно реакции вполне однозначны. Например, при рассмотрении
кратковременных эффектов во всех случаях при увеличении ∆w наблюдается уменьшение ci,
при увеличении ca –– увеличение ci и т.д.; при долговременном увеличении интенсивности
освещения I во всех случаях поднимается плато световой кривой, при увеличении ca началь-
ный наклон углекислотной кривой уменьшается и т.д.
Такой однозначности реакций нет при действии водного дефицита или какого-либо дру-
гого фактора, требующего изменений с точки зрения осмотической адаптации –– прежде всего
засоления. Можно наблюдать самые разнообразные реакции для тех же воспроизводимых ха-
рактеристик, которые изменяются вполне однозначно при действии перечисленных выше фак-
торов. Разнообразие демонстрируют даже данные совсем небольшой совокупности цитируе-
мых ниже работ, а не сотен работ, описывающих гораздо более однозначную реакцию для
многочисленных факторов, рассмотренных в пп.5–6 и приложении С.
А именно, при действии водного дефицита <определяемого через изменение водного по-
тенциала листа ψl> и/или засоления значение ci может уменьшаться (весьма яркий пример ––
данные [Brodribb, 1996]), оставаться примерно постоянным [Mooney et al., 1977], увеличивать-
ся [Rensburg, Kruger, 1993]. Начальный наклон углекислотной кривой может падать [Mooney et
al., 1977; Matthews, Boyer, 1984; Ball, Farquhar, 1984a; Rensburg, Kruger, 1993], оставаться не-
изменным [Brodribb, 1996; Ball, Farquhar, 1984b] (или даже несколько увеличиваться
[Matthews, Boyer, 1984]), причем даже по данным для одного и того же вида [Matthews, Boyer,
1984; Ball, Farquhar, 1984a,b]. Квантовый выход фотосинтеза и эффективность электронного
транспорта могут уменьшаться или оставаться постоянными [Mooney et al., 1977; Matthews,
Boyer, 1984]. Значение УКП может увеличиваться [Rensburg, Kruger, 1993] или оставаться по-
стоянным (причем последнее возможно даже в условиях, когда уже явно выражена деструкция
физиологических процессов (как для Aegiceras corniculatum по данным [Ball, Farquhar, 1984a]).
В некоторых случаях при действии осмотических ограничений происходит явное изменение
параметров сопряжения при интеграции световых и темновых процессов фотосинтеза –– уси-
ливается жесткость углекислотных кривых [Ball, Farquhar, 1984a].
Область значений ψl, в которой происходят наиболее существенные изменения текущего
значения скорости ассимиляции и перечисленных характеристик, весьма различается у разных
объектов и даже одного и того же объекта в зависимости от предадаптации [Matthews, Boyer,
1984 (см. Рис.6); Plaut, Federman, 1991; Mooney et al., 1977]. Причем в то же самое время в не-
которых случаях наблюдается сходство реакций для весьма различающихся объектов с точки
зрения актуальности для них проблемы осмотической адаптации. Например, ниже дано срав-
74
нение данных для таких разных видов, как вид Larrea divaricata при экстремальных условиях
водного дефицита в естественном для этого вида местообитании в Долине смерти (Калифор-
ния, США) и культурный вид Heliathus annuus (подсолнечник), выращиваемый в условиях
гораздо более благоприятного водного режима.
Интересно, что все это разнообразие реакций, по-видимому, полностью согласуется с
предшествующими этапами оптимизационного описания, если выражать ограничения при ос-
мотической адаптации <в связи с необходимостью обеспечить неотрицательный водный ба-
ланс листа> соотношением вида (14) или любым аналогичным способом, подразумевающим
критическую зависимость от водного потенциала. В этом смысле проводимый ниже анализ
имеющихся фактов подтверждает, прежде всего, правильность предшествующего оптимиза-
ционного описания, а не гипотезу о <«вытесняющем» или аналогичном с точки зрения коли-
чественного описания деструктивном кооперативном неспецифическом> механизме осмоти-
ческой адаптации. Проводимый анализ не является прямым доказательством точности форму-
лы (14) как способа описания ограничений в связи с осмотической адаптацией.
Вывод о согласовании с предшествующими этапами оптимизационного описания следу-
ет из применения полученных оптимизационных соотношений в естественной интерпретации,
рассматривая действие водного дефицита как влияние ψl на параметры в оптимизационных
соотношениях. В первую очередь представляют интерес не слишком кратковременные воздей-
ствия (таковыми являются засуха и засоление при их естественном развитии, а кратковремен-
ная реакция на водный дефицит в воздухе рассмотрена далее в п.7.1.3). Тогда успевает про-
изойти перестройка всех процессов, и можно пользоваться оптимизационными соотношения-
ми в полном объеме, в частности, уравнениями (8) –– (9) для описания интеграции световых и
темновых реакций фотосинтеза. Для оценки ci вместо более общего уравнения (10) удобно
использовать более простое выражение (С.8), которое было получено в модели устьичной ре-
гуляции фотосинтеза.
Из оптимизационных соотношений следует:
– пока ограничения в связи с водным дефицитом выражены слабо (рабочая точка нахо-
дится на верхнем плато кривой A(ψl) –– см. рис.6), водный дефицит означает увеличение
удельных затрат αw при неизменных затратах на фотосинтез (αc и αJ); оптимизационное со-
отношение в упрощенной форме (С.8) или в более общем (10) предсказывает действительно
наблюдаемое во всех случаях (пока водный дефицит не критичен) уменьшение ci;
– с вытеснением фотосинтеза или его деструкцией в результате неспецифического коо-
перативно воздействия <предсказываемых уравнением (14) или любым аналогичным> удель-
ные затраты на фотосинтез возрастают, причем особенно быстро при приближении к критиче-
скому значению ψl. В пределе приближения к критическому значению ψl удельные затраты на
фотосинтез формально становятся сколь угодно велики, тогда как удельные затраты αw воз-
растают, но не столь быстро и в любом случае конечны. Отсюда вытекает необходимость в
смене тенденции с убывания ci к возрастанию, которая также обычно проявляется во всех слу-
чаях на последних стадиях развития водного дефицита перед окончательной деструкцией фо-
тосинтетического аппарата и гибелью листа. Тенденция к росту ci должна быть выражена тем
более, если с усилением водного дефицита (при все более отрицательных ψl) увеличивается
УКП <как, например, наблюдаем для табака по данным [Rensburg, Kruger, 1993]>.
75
0 100 200 300
Сi, мкл/л
0
10
20
30
Скорость ассимляции, мкмоль СО2 /м2 с
Рисунок 7 (сохранение наклона углекислотной кривой –– технически модифицированный
рисунок из работы [Brodribb, 1996]). Связь скорости ассимиляции (A) и концентрации CO
2
в
межклетниках (c
i
) для Podocarpus lawrencii: о –– в случае действия водного дефицита при под-
держании постоянной внешней [CO
2
] (c
a
= 360 мкл/л, стрелки показывают тенденцию измене-
ния c
i
с усилением водного дефицита); ▼ –– в благоприятных условиях при изменении c
a
(ко-
эффицент корреляции для указанной линейной регрессии r = 0.935).
Данные в процитированных выше работах полностью подтверждают описанную основ-
ную тенденцию. Наиболее ярко все <выделенные> фазы реакции на водный дефицит демонст-
рирует рисунок 7. Есть основания ожидать, что для этих и других случаев удастся установить
не только качественное, но и количественное соответствие оптимизационному описанию на
предшествующих этапах, т.к. такое согласование прямо или косвенно подтверждает сопостав-
ление различного рода данных, приведенных в этих работах.
Так, в работе [Matthews, Boyer, 1984] для растений подсолнечника, адаптированных и не-
адаптированных к водному дефициту, в первом случае (для адаптированных) эффективность
электронного транспорта не изменялась существенно в значительно более широком диапазоне
ψl, чем для неадаптированных (примерно до –15 бар в сравнении с –7 бар). Поэтому нет осно-
ваний считать, что в этом диапазоне происходило изменение удельных затрат на фотосинтез.
Таким образом, с оптимизационными соотношениями полностью согласуется тот факт, что
при значении ψl из этого диапазона наклон углекислотной кривой и уровень ее плато не
уменьшаются.
Во втором случае (при исследовании реакции неадаптированных растений) в том же диа-
пазоне ψl наблюдалось падение эффективности электронного транспорта, т.е. удельные затра-
ты на фотосинтез (прежде всего, αJ) неизбежно возрастали. При этом разовые затраты Q0 сле-
дует считать постоянными. Статические затраты q0 к моменту достижения листом зрелости
уже в основном сделаны и не зависят от последующего воздействия. Нет также оснований
считать, что изменилось время функциональной активности листа τ. В частности, нет кинети-
ческого эффекта типа накопления соли в листе и ее быстро усиливающегося деструктивного
влияния на фотосинтез как при действии даже слабого засоления на неустойчивые к соли ви-
ды. Таким образом, действие водного дефицита следует интерпретировать как увеличение от-
ношения αJ/Q0, при котором целесообразно наблюдаемое уменьшение всех скоростей –– мак-
76
симальной скорости Jm (понижения плато углекислотной кривой), скорости карбоксилирова-
ния vc (о карбоксилировании и изменении vm будет сказано ниже отдельно) и действующей
скорости A. Значительное отличие в сравнении с адаптированными растениями вполне объяс-
нимо также и тем, что для неадаптированных растений механизм осмотической адаптации
предположительно менее совершенен, что выражается в том, что он <в соответствии с уравне-
нием (14) или ему подобным> сильнее «вытесняет» все остальные механизмы, т.е. прежде все-
го, фотосинтез.
Аналогично по данным [Mooney et al., 1977] для Larrea divaricata при выращивании в ла-
боратории квантовый выход фотосинтеза падает. Для сравнения гораздо более быстрое паде-
ние квантового выхода наблюдалось и в случае неадаптированных растений подсолнечника <в
рассматриваемом при сопоставлении с адаптированным растениями диапазоне значений ψl>.
Поэтому наблюдаемое в этом случае падение Jm, vc и A также находится в полном соответст-
вии с экономической целесообразностью. Интересно, что для этого же вида в естественном
местообитании квантовый выход не падает даже при ψl = –50 бар. Исходя из этих данных,
можно в соответствии с той же экономической целесообразностью предположить (данные в
цитируемой работе проверить такое предположение не позволяют), что действующее значение
A при том же ψl (пока квантовый выход не изменяется) будет изменяться очень слабо как ре-
зультат первой фазы реакции на водный дефицит –– уменьшения ci.
Применительно к первой фазе реакции на водный дефицит можно сделать еще одно общее
предположение. А именно, в рамках неспецифического кооперативного действия в связи с
<осмотической> адаптацией в водному дефициту можно ожидать изменения соотношения
между удельными затратами на карбоксилирование и удельными затратами на другие про-
цессы при фотосинтезе. В благоприятных условиях, когда водный дефицит не проявляется,
выделения с экономической точки зрения одной реакции карбоксилирования из множества
других вполне обосновано. Осуществление этой одной реакции <карбоксилирования> срав-
нимо по затратам со всеми реакциями электронного транспорта –– в силу того, что соответ-
ствующий фермент РДФК имеет большую М.м. 550 кДа (на порядок больше характерной 30
кДа) и при этом низкую производительность (на порядок меньше характерной). Но с осмо-
тической точки зрения оснований для аналогичного особого выделения карбоксилирования
нет –– вклад низкомолекулярных участников этой реакции значителен не более, чем для
любой другой ферментативной реакции. Вклад высокомолекулярных образований и самой
РДФК в осмотическое давление тем более незначителен. Наличие РДФК и других крупных
макромолекулярных агрегатов (полисом, клеточных стенок и т.п.) следует представить как
уменьшение эффективного объема водной фазы путем вычитания объема этих агрегатов.
Таким образом, с усилением водного дефицита и усилением значимости осмотического
фактора экономическая значимость карбоксилирования в сравнении с другими реакциями и
77
процессами предположительно будет уменьшаться. В терминах удельных затрат такой гипоте-
тический результат действия водного дефицита следует определить как более медленное уве-
личение затрат на карбоксилирование в сравнении с увеличением удельных затратами на
обеспечение других этапов фотосинтеза.
Если предположение о дифференциальном влиянии водного дефицита на карбоксилиро-
вание в сравнении с другими процессами справедливо, то экономически целесообразную ре-
акцию на первую фазу водного дефицита должно выражать примерно пропорциональное
уменьшение максимальных скоростей всех основных этапов фотосинтеза, но не максимальной
скорости карбоксилирования vm. Уменьшение последней в сравнении с другими максималь-
ными скоростями должно происходить значительно медленнее или отсутствовать.
Именно такое ожидаемое изменение соотношения максимальных скоростей будет, во-
первых, соответствовать предполагаемому характеру изменения соотношения удельных за-
трат. Во-вторых, тем самым будет учтено также, что при первой фазе реакции на водный де-
фицит целесообразно связанное с необходимостью большей экономии водных ресурсов
уменьшение ci, которое и при постоянном значении vm означает уменьшение действующей
скорости карбоксилирования vc. Согласование скорости карбоксилирования с уменьшением
скоростей других процессов (электронного транспорта, совокупности реакций цикла Кальви-
на, активного или активированного транспорта СО2 к местам карбоксилирования и ассимиля-
тов к местам загрузки) отчасти происходит в результате уменьшения ci и без уменьшения vm.
Если же согласование скоростей произойдет другим способом, например, за счет умень-
шения значения vm при постоянном значении ci, то требуемый более экономичный расход
воды достигнут не будет, и не будет рационально использовано предполагаемое относитель-
ное уменьшение экономической значимости карбоксилирования.
Таким образом, в модели интеграции карбоксилирования с другими процессами можно
считать, что необходимость уменьшения максимальных скоростей при осмотической адапта-
ции <описываемая формулой (14) как изменение общего масштаба процессов fm> относится
не к карбоксилированию, а к другим процессам. Для интеграции процессов при фотосинтезе
такое утверждение конкретизирует необходимость в уменьшении общего масштаба происхо-
дящих процессов при любом способе осмотической адаптации –– общую тенденцию к вытес-
нению механизмом осмотической адаптации всех других механизмов из активного объема,
выражаемую формулой (14) или другой аналогичной.
В этом смысле выражаемые формулой (14) ограничения относятся к значениям дейст-
вующих и максимальных скоростей в расчете на единицу активно функционирующего объема.
Однако расчет на единицу активно функционирующего объема очень сложно осуществить
практически, т.к. активная зона занимает заведомо меньшую часть объема фотосинтезирую-
щей клетки (большую часть занимает относительно инертная вакуоль), а лист, в свою очередь,
имеет сложную архитектуру как совокупность клеток.
Если бы можно было измерять скорости в расчете на единицу активно функционирую-
щего объема, то тогда можно было бы путем прямых измерений устанавливать ограничения
при осмотической адаптации, выражая их в виде формулы, подобной формуле (14). В частно-
сти, правильность формулы (14) можно было бы проверить непосредственно, поскольку все
величины, входящие в (14), определены как независимые и в принципе их можно измерить
независимо. Величины fm и ψ можно было бы измерить как текущие значения скорости инте-
ресующего физиологического процесса и водного потенциала, fm0 –– как предельное значение
fm в отсутствии осмотических ограничений, πкрит –– как предельное <при котором fm падает
до нуля> для используемого осмотика или композиции осмотиков.
Однако скорости в расчете на единицу активного объема практически недоступны для
прямых измерений, зато удается точно измерить значения скоростей в расчете на единицу
проективной площади листа, а поэтому важно понимать, как ограничение (14) можно связать
со значениями скоростей, измеряемыми на единицу площади.
Прежде всего, важно понимать, что уменьшение общего масштаба скоростей, рассчитанных
на единицу площади, не является <физически> неизбежным следствием вытеснения меха-
78
низмом осмотической адаптации других механизмов из активного объема. Физически воз-
можно даже увеличение масштаба скоростей, рассчитанных на единицу площади, за счет
многократного увеличения активного объема, приходящегося на единицу площади листа.
Причем при таком многократном увеличении в принципе не является неизбежным увеличе-
ние измеряемой толщины листа, поскольку мал вклад активного объема в объем клетки, а в
силу значительного объема воздушных полостей в листе суммарный объем клеток сущест-
венно меньше объема листа, определяемого как произведение проективной площади листа
на его толщину.
В терминах экономических характеристик уменьшение эффективности в единице активного
объема можно компенсировать за счет многократного увеличения затрат на единицу пло-
щади. Иными словами, падение эффективности в расчете на фотосинтезирующую единицу
компенсирует <более быстрое> увеличение количества отдельных составляющих в составе
фотосинтезирующих единиц и количества фотосинтезирующих единиц в целом на единицу
площади.
Но такое компенсирующее изменение было бы заметно как многократное увеличение со-
держания белков и других составляющих фотосинтезирующих единиц на единицу площади.
Обычно такого увеличения не наблюдают и это вполне объяснимо.
Экономическая целесообразность аналогичной, относительно консервативной, реакции
уже обсуждалась применительно к реакции на изменение интенсивности освещения при вы-
ращивании (см. п.6.3.3). Наблюдаемая слабая восприимчивость максимальных скоростей к
интенсивности освещения I <изменение масштаба скоростей медленнее, чем пропорционально
корню квадратному из I> полностью соответствует целесообразной в соответствии с оптими-
зационными соотношениями (8) –– (9).
Таким образом, вполне естественно распространить применение формулы (14) или ана-
логичной ей для расчета скоростей на единицу площади. В случае применения по отношению
к скоростям, рассчитываемым на единицу площади, формула (14) приобретает смысл оптими-
зационного соотношения как выражение представления об экономической целесообразности.
Как способ выражения ограничений для скоростей, рассчитываемых на единицу площа-
ди, формула (14) <а следовательно, и любое основанное на ней утверждение> имеет прибли-
женный характер с учетом того, что происходит некоторое изменение количества составляю-
щих фотосинтезирующих единиц на единицу площади. Как правило, регистрируют изменения
содержания белка и т.п. изменения количеств составляющих, измеряемые десятками процен-
тов. Приближенные утверждения, получаемые при применении (14), можно уточнить при на-
личии соответствующих данных.
В частности, сделанный выше вывод о постоянстве значения vm для первой фазы реак-
ции на водный дефицит будет справедлив при обычном способе расчета величины vm на еди-
ницу площади. Наиболее яркая иллюстрация постоянства начального наклона углекислотной
кривой в широком диапазоне изменения функционального значения ci (сдвига рабочей точки
79
при уменьшении водного потенциала листа до –25 бар) представлена на рисунке 7. Описывае-
мая реакция постоянства начального наклона почти вплоть до сдвига рабочей точки к УКП ––
это крайний случай. Уменьшение начального наклона (которое вполне целесообразно, если
происходит увеличение удельных затрат на карбоксилирование) обычно наблюдают задолго
до приближения к УКП.
Другой универсальный вывод следует из (14) при описании всех фаз реакции на водный
дефицит, если распространить применение этой формулы по отношению к скоростям, рассчи-
тываемым на единицу площади. Это вывод о линейной связи водного потенциала листа с об-
щим масштабом наблюдаемых действующих и максимальных скоростей.
Интересно, что многочисленные данные подтверждают такой вывод. В частности, вывод
о приближенно линейной связи действующих скоростей и ψl позволяют проверить данные,
представленные в некоторых из цитируемых выше работ [Mooney et al., 1977; Plaut, Federman,
1991; Rensburg, Kruger, 1993; Brodribb, 1996]. Приближенная линейность действительно на-
блюдается, причем в последней из этих работ подтверждение линейной связи дано в своеоб-
разной косвенной форме. А именно, с одной стороны, продемонстрировано сохранение накло-
на углекислотной кривой в широком диапазоне значений и последующее увеличение ci с уси-
ление водного дефицита (Рис.7). Таким образом, минимально функциональное значение ci
имеет смысл нижней граничной точки на линейном участке углекислотной кривой с сохра-
няющимся наклоном. С другой стороны, неявно показано также подобие углекислотных кри-
вых у исследованных видов. О подобии говорит приводимый выбор близких наблюдаемых
рабочих значений ci в благоприятных условиях и близость значений УКП для всех C3-видов в
этих условиях. С учетом подобия минимальное функциональное ci линейно связано с дейст-
вующей скоростью фотосинтеза. Следовательно, косвенным подтверждением линейной связи
между действующей скоростью фотосинтеза и ψl является полученная линейная связь мини-
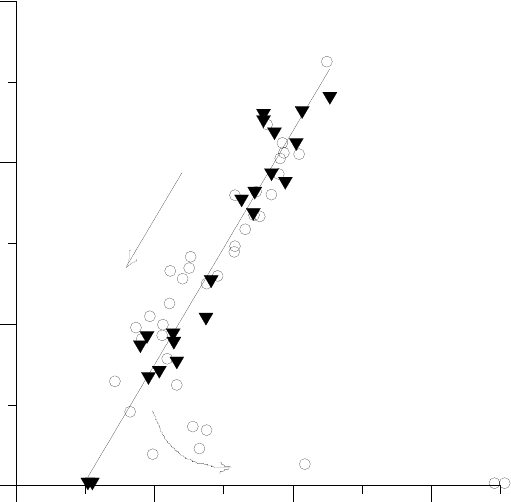
мально функционального ci и ψl (Рис.8).
В общем случае разнообразие реакций на водный дефицит <в связи с фотосинтезом> оп-
ределяют особенности осмотических систем <фактор a>, критические значения ψl <фактор
nкрит> для совокупности физиологических процессов и каждого из процессов по отдельности
<а подробнее, зависимость удельных затрат αc(ψl) и αJ(ψl)>, а также варианты жизненных
стратегий <фактор τ и связанная с ним величина Q0>. Для любого объекта перечисленные ха-
рактеристики следует считать заданными (их можно установить независимо), а зная их, можно
предсказать экономически целесообразную реакцию на водный дефицит для конкретного объ-
екта.

80
-4.0 -3.0 -2.0
Водный потенциал листа, МПа
0.10
0.20
0.30
0.40
(Ca/Ci)min
3
4
9
6
8
7
2
5
1
Рисунок 8. <Линейная> связь минимального функционального c
i
(представленного в виде
отношения c
i
/c
a
, где c
a
близко к обычному атмосферному значению) и водного потенциала при
этом c
i
для 9 видов хвойных (показана линейная регрессия, коэффициент корреляции 0.976): 1
–– Acmopyle pancheri, 2 –– Athrotaxis selaginoides, 3 –– Actinostrobus acuminatus, 4 –– Callitris
rhomboidea, 5 –– Dacrycarpus dacrydioides, 6 –– Diselma archerii, 7 –– Lagarostrobos franklinii, 8 –
– Microstrobos niphophilus, 9 –– Podocarpus lawrencii.
Полезными оказываются не только прямые измерения входящих в оптимизационные со-
отношения (8) –– (10) величин <в параметрической зависимости от ψl> (а величины αc(ψl) и
αJ(ψl) достаточно сложно измерить непосредственно), но и измерения других обычно наблю-
даемых <более доступных> величин. Некоторые оценки типа неравенств по отношению к
удельным затратам следуют из измерения эффективности электронного транспорта и кванто-
вого выхода фотосинтеза. Интерпретация данных этого рода уже была использована выше при
сравнении данных для Larrea divaricata и Heliathus annuus. Прямым показанием увеличения
затрат на карбоксилирование является рост УКП с усилением водного дефицита. Косвенным
индикатором увеличения затрат на карбоксилирование является уменьшение наблюдаемой
производительности на единицу массы РДФК –– естественно предположить примерную про-
порциональность между этими величинами. Еще более косвенно об увеличении <динамиче-
ских> затрат αc и αJ свидетельствует увеличение общей массы белка на единицу площади,
еще более косвенно –– сухого веса, хлорофилла и т.п. Прямое показание увеличения затрат Q0
–– сокращение времени функциональной активности листа τ, например, в результате деструк-
ции при водном дефиците или накопления соли в листьях неустойчивых к соли растений.
Действующее значение водного потенциала является важнейшей характеристикой <интен-
сивно> транспирирующей ткани в силу предполагаемой прямой связи <в трактовке линей-
ного вытеснения или шире –– линейного эффекта кооперативного действия> водного по-
тенциала листа со скоростями физиологических процессов. Наблюдение за изменением