Реферат - изменчивость микроорганизмов
Подождите немного. Документ загружается.

Министерство здравоохранения Украины
Запорожский государственный медицинский университет
Кафедра микробиологии, вирусологии и иммунологии
Реферат на тему:
Изменчивость
микроорганизмов
Выполнила студентка 13 группы медицинского
факультета Кривокрисенко М.Ю.
Проверила: асс.Моргунцова С.А,
Зав.кафедрой д .м.н. Силина Т.Н.
Запорожье 2009г

План:
1.Изменчивость микроорганизмов.
2.Фенотипическая изменчивость
3.Генотипическая изменчивость
4. Изменчивость фагов и изменчивость микроорганизмов под влиянием фагов
5. Использование генетики микроорганизмов
6.Литература
2
ИЗМЕНЧИВОСТЬ БАКТЕРИЙ
Наследственность консервативна, она обуславливает стабильность вида микроорганизмов,
напротив, изменчивость является выражением способности вида приспосабливаться к постоянно
меняющимся условиям его обитания. Наследственность и изменчивость неразрывно связаны
между собой и размножением микроорганизмов. В популяции бактерий всегда появляются
клетки, которые могут менять свои свойства. Если изменение признаков под влиянием факторов
различного порядка, связаны с генотипом бактерий, то эти изменения передаются по наследству
и могут быть положительно расценены естественным отбором. Новые признаки,
обеспечивающие селективное преимущество, закрепляются естественным отбором, меняется
генотип вида, осуществляется процесс эволюции.
У микроорганизмов различают фенотипическую (модификационную, ненаследственную) и
генотипическую (наследственную) изменчивость.
3
Фенотипическая изменчивость"
Фенотипическая изменчивость является не наследуемым типом изменчивости, т. е. это различия
между микроорганизмами, одинаковыми по генотипу. Эта изменчивость возникает в результате
постоянного воздействия на клетку изменяющихся факторов среды обитания. Сходные по
генотипу, микроорганизмы могут существенно различаться по фенотипу, т. е. по способу
проявления наследственных признаков.
На формирование фенотипа существенное влияние оказывают факторы внешней среды.
Известно, что генотипически идентичные организмы в различных условиях существования в
определенной степени различаются по своим признакам. Например, изменение содержания
жира в молоке животных или массы тела в зависимости от их кормления, изменение количества
эритроцитов в крови в зависимости от порциального давления кислорода.
В отличает от особей высший организмов, у которых исследуются признаки каждой особи, у
микроорганизмов изучают не признаки одной клетки, а всей культуры, которая включает
миллиарды бактерий. Культуры микробов, выращенные на питательной среде, отличаются
характером роста, физиологическими и биохимическими признаками. К морфологическим
признакам относят окраску, размер, форму, наличие жгутиков, капсул, спор и т. д. К
физиологическим признакам культур относятся способность расти при определенной
температуре, устойчивость к химическим веществам, облучению, антибиотикам, фагам,
различным ядам.
3
Примером модификационной изменчивости у микроорганизмов может быть образование
различных типов адгезинов у гонококка, необходимых для колонизации им кишечника. В
качестве примера, можно привести увеличение сальмонелл при добавлении к питательной среде
стрептомицина. При переносе таких сальмонелл в питательную среду без стрептомицина
бактериальные клетки приобретают типичную для вида величину.
Модификации представляют собой изменения, которые поддерживаются пока действует
неблагоприятный фактор. Так, образование L-форм бактерий, лишенных клеточной стенки,
происходит под влиянием химиотерапевтических веществ (пенициллина, стрептомицина и т. д.).
при снятии действия антибиотиков на культуру бактерий происходит реверсия
микроорганизмов в исходные формы. Фенотипическое проявление признака под влиянием
условий внешней среды возможно в определенных пределах, называемых нормой реакции,
которая допустима генотипом организмов. Некоторые признаки характеризуются широкой
нормой реакции. В основном, это количественные признаки (масса микробной клетки, ее
величина, пигментация колоний).
Фенотипическое проявление информации, заключенной в генотипе, характеризуется
показателями пенентрантности и экспрессивности. Пенетрантность отражает частоту
фенотипического проявления имеющейся в генотипе информации, а экспрессивность
характеризует степень выраженности признака.
Различают длительную модификацию, которая проявляется в течение нескольких поколений и
кратковременную, при которой изменения исчезают при исчезновении действующего фактора
внешней среды.
Генотипическая изменчивость
Генотипическая изменчивость связана с изменением генотипа бактерий. В основе
генотипической изменчивости лежат мутации и рекомбинации.
Мутации (от латинского mutatio - изменение) - это изменения3 структуры ДНК (качественные
или количественные), которые возникают под влиянием эндогенных или экзогенных факторов и
проявляются наследственно закрепленным изменением одного или многих признаков. В
природе мутации возникают без участия экспериментатора и называются спонтанными, а
мутации, контролируемые экспериментатором, называются индуцированными. Бактерии с
измененными признаками называют мутантами. Спонтанные мутации возникают под влиянием
неизвестных причин и лежат в основе эволюции микроорганизмов. Факторы, вызывающие
мутации, называются мутагенами. Различают физические, химические и биологические
мутагены.
К физическим мутагенам относятся такие факторы, как температура, радиация,
ультрафиолетовые лучи, ионизирующие излучения и др.
К химическим мутагенам принадлежат многочисленные химические соединения и вещества,
которые могут изменять структуру генов, взаимодействуя с ДНК бактериальной клетки.
4
Биологическими мутагенами являются бактериофаги и продукты жизнедеятельности клеток,
которые накапливаются в питательной среде в результате размножения и роста бактерий.
По широте изменений генома бактерий мутации делят на генные - изменения регистрируют в
пределах одного гена, хромосомные - в группе генов, точковые - в одном триплете.
В зависимости от взаимодействия мутагенов на нуклеотид бактериальной клетки или ее
плазмиды, мутации делят на нуклеоидные и плазмидные.
По направлению выделяют прямые и обратные мутации. Прямые - это изменения генов
бактерий, выделенных из естественной среды обитания. Обратные мутации - это возврат от
измененного типа бактерий к естественному типу.
По фенотипическому проявлению различают нейтральные, условно-летальные и летальные
мутации.
Нейтральные мутации фенотипически не проявляются. Условно-летальные мутации ведут к
изменению, но не к исчезновению функциональной активности фермента. Летальные - это
мутации, ведущие к полной потери способности клетки синтезировать жизненно необходимые
ферменты, что приводит к ее гибели.
Мутации фенотипически проявляются изменением морфологических, биохимических,
вирулентных и других свойств.
Диссоциация - это особый, присущий только бактериям вариант изменчивости, при котором
происходит культуральная изменчивость, т. е. расщепление вида и возникновение при росте на
плотной питательной среде двух основных типов колоний: S-форма - гладкие (от английского
smooth - гладкий) и R-форма (от английского rough - шероховатый) - шероховатые. Между
этими формами имеются и переходные М-, О-, Д-формы.
Микроорганизмы из колоний в S-форме обладают хорошо выраженными антигенными и
вирулентными свойствами и, напротив, у бактерий из колоний в R-форме эти свойства
выражены слабо. Однако, не всегда S-форма микробов является свидетельством их
вирулентности. Например, возбудитель сибирской язвы, туберкулеза, чумы вирулентны в R-
форме.
В основе диссоциации лежат мутации, спонтанно возникающие в естественной среде обитания
микробов или же при культивировании их на искусственных питательных средах.
Диссоциация имеет большое значение для микроорганизмов, так как они, благодаря этому
явлению, получают селективное преимущество, обеспечивающее их существование в организме
животных и человека, а также во внешней среде. Известно, что S-формы более устойчивы к
фагоцитозу, R-формы - к факторам естественной среды обитания.
Геном бактерий способен к репарации. Репарация - это процесс восстановления структуры
поврежденной ДНК, который обеспечивается многочисленными ферментами, определяющими
5
состояние этой кислоты. Например, фоторепарация зависит от фотолиаз. Эти ферменты
активизируются при образовании тиминовых димеров в ДНК под воздействием
ультрофиолетового облучения и деполизируют эти димеры до исходных мономеров.
Наибольшее значение в жизнедеятельности микроорганизмов имеет SOS-репарация или SOS-
ответ. SOS-ответ - это реакция микробных клеток на прекращение синтеза нуклеиновых кислот
в связи с повреждением ДНК, голоданием клетки, воздействием продуктов метаболизма и т. д.
SOS-ответ возникает при критическом состоянии клетки, на грани ее гибели, как реакция
направленная на восстановление жизнедеятельности клетки. Например, результатом SOS-ответа
у E. Coli является синтез около 25 белков, имеющих непосредственное отношение к репарации,
рекомбинации и синтезу ДНК. SOS-ответ у микроорганизмов контролируется SOS-областью.
Обычно гены этой области находятся в неактивном состоянии и активизируются лишь в
критические для жизни клетки моменты. SOS-репарация обеспечивает развитие микробной
популяции в целом и ее адаптацию к изменившимся внешним условиям.
Кроме мутаций у бактерий известны рекомбинационная изменчивость. Рекомбинация - это
передача генетического материала от клетки-донора с одним генотипом к клетке-реципиенту с
другим генотипом. В результате такой передачи образуются рекомбинанты - т. е. бактерии,
обладающие свойствами обоих родителей. Рекомбинация является важнейшим фактором
эволюции, т. к. между разными особями происходит обмен генетической информацией, что
повышает уровень их приспосабливаемости к различным внешним факторам окружающей
среды. Рекомбинации могут наблюдаться на уровне любых живых организмов - от прокариот до
высших эукариот.
Различают следующие способы рекомбинационной (комбинативной) изменчивости:
трансформация, трансдукция, конъюгация.
Трансформация (от латинского transformo - превращать, преобразовывать) - изменение генома
бактерий - реципиента, в результате поглощения из среды свободного фрагмента ДНК клетки-
донора.
Впервые явление трансформации начал изучать Ф. Гриффитс (1928), используя в опытах
культуры пневмококков. Эти микроорганизмы способны к диссоциации и образуют на плотной
питательной среде колонии в S-форме и R-форме. Микроорганизмы образующие S-формы
колоний капсульные,3 они патогенны для белых мышей. Бактерии, формирующие на агаре R-
формы колоний бескапсульные, не патогенные для мышей. Фактором патогенности у
пневмококков является капсула, что было учтено Ф. Гриффитсом при проведении опытов. Он
ввел мышам вместе две культуры пневмококков: одну - непатогенную бескапсульную (R-
штамм), а вторую - патогенную с капсулой (S-штамм), но обезвреженную нагреванием. Мыши,
получившие смесь упомянутых культур пали. Из крови павших мышей была получена культура,
микроорганизмы которой имели капсулу и обладали патогенностью. Контрольные3
эксперименты продемонстрировали, что введение мышам по отдельности живых пневмококков
бескапсульных и убитых нагреванием не приводит к гибели животных. Ученый сделал вывод,
что непатогенные клетки R-штамма могут3 рансформироваться в патогенные пневмококки,
обладающие капсулой.
6
Грачевой (1946) был получен вариант кишечной палочки с некоторыми свойствами
характерными для сальмонелл. Она3 культивировала E. Coli на среде, к которой добавлялась
убитая культура сальмонелл.
В результате многочисленных экспериментов было установлено, что путем трансформации
могут быть перенесены различные признаки: синтез капсульного полисахарида, синтез
различных ферментов, устойчивость к антибиотикам и т. д.
Было обнаружено, что трансформация имеет место чаще в пределах одного вида, но может
наблюдаться и между разными видами. В процессе трансформации участвуют две
бактериальные клетки: донор и реципиент.
О. Эвери, К. Мак-Леод, М. Мак-Карти (1944) установили, что трансформирующим фактором
является ДНК. По их мнению, трансформация представляет собой поглощение изолированной
ДНК бактерии донора клетками бактерии реципиента.
Трансформация - сложный биологический процесс, который протекает поэтапно. Первая стадия
этого процесса заключается в адсорбции трансформирующей ДНК на поверхности микробной
клетки. Вторая - проникновение ДНК через определенные рецепторные участки стенки
бактерии-реципиента при помощи специальных белков внутрь клетки. Третья стадия
представляет собой спаривание части ДНК донора с ДНК реципиента, четвертая - включение в
ДНК реципиента одной из цепей трансформирующего элемента. И пятая - изменение
нуклеотида клетки-реципиента в ходе ее последовательных делений. Способность бактерий
реципиентов к трансформации была названа компентентностью. Компентентность определяется
физиологическим состоянием клетки-реципиента к периодам клеточного цикла.
Трансдукция (от латинского transductio - перенос) - перенос генов из одной бактериальной
клетки в другую при помощи бактериофага. Явление трансдукции впервые установили Н.
Циндлер и ДЖ. Ледербер (1952). Для исследований они использовали патогенные для белых
мышей два штамма S. typhimurium (22 A и 2A). Штамм 22 А - ауксотрофный, не способный
синтезировать триптофан (Т
-
), штамм 2А - способный к синтезу триптофана (Т
+
). В опытах
исследователи использовали U-образную трубку, разделенные на изгибе бактериальным
фильтром. В одно колено этой трубки с питательной средой засевали бактерии штамма 22 А, в
другое - штамма 2А. Опыты показали, что штамм 22 А был лизогенен по фагу Р-22. Этот фаг из
лизогенной культуры проходил через бактериальный фильтр, лизировал бактерии штамма 2А,
присоединял при этом его генетический материал. Затем фаг возвращался обратно и передавал
генетический материал штамма 2А штамму 22А, который приобретал способность
синтезировать триптофан.
Явление трансдукции установлено не только у сальмонелл, но и у кишечной палочки и
актиномицетов. У бактерий наблюдается трансдукция одного, реже двух и весьма редко трех
сцепленных генов.
Различают следующие виды трансдукции: общую (неспецифическую), специфическую и
абортивную.
7
Общая трансдукция характеризуется тем, что фаг играет роль переносчика генетического
материала бактерий, т. е.3 передает в клетку-реципиент любой ген донорской клетки. Сам фаг в
нуклеоид реципиента не встраивается и лизогении бактериальной культуры не происходит.
Один и тот же фаг может служить трансдуктором различных признаков: ферментативной
активности, устойчивости к лекарственным веществам, подвижности, вирулентности и др.
Специфическая трансдукция заключается в том, что бактериофаг переносит от клетки-донора в
клетку-реципиента строго определенные гены и встраивает их в определенные участки
реципиента. Бактериофаг может встраиваться в нуклеоид клетки-реципиента. Клетки бактерий,
имеющие в своей хромосоме профаг, называют мезогенными, а явление совместного
существования ДНК бактерий и профага называется мезогенным.
Абортивная трансдукция характеризуется тем, что фрагмент ДНК донора, перенесенный в
клетку реципиента не включается в ее нуклеоид, а может сохраняться в цитоплазме клетки.
Клетка при этом не подвергается лизису, но при делении ее перенесенный новый признак
постепенно исчезает у ее потомства.
Конъюгация (от латинского conjugatio - контактирование) - перенос генетического материала от
одной бактериальной клетки (донора) к другой (реципиенту) при непосредственном контакте
этих клеток. Явление3 конъюгации открыли Дж. Ледерберг и Э. Татуш (1946).
Ученые взяли два ауксотрофных мутантных штамма E. Coli к-12: один не способный
синтезировать треонин и лейцин (Thr-Leu
-
), другой - метионин и биотин (Met-Bio
-
) и
выращивали их вместе в течение 12 часов на полноценной питательной среде. Затем выросшую
культуру отцентрифугировали и отмыли от полноценной питательной среды и засеяли на
минимальную питательную среду.
На этой среде без метионина, биотина, треонина и лейцина появились прототрофные колонии
Met
+
, Bio
+
, Thr
+
, Leu
+
. Опытным путем ученые установили, что ни трансформации, ни
трансдукции в данном случае не наблюдалось. Был сделан вывод о происхождении
рекомбинантных геномов в результате непосредственного контакта родительских клеток.
Микрофотографии конъюгирующих клеток явились доказательством того, что между ними
образуется цитоплазматический мостик.
В 1952 году Хейтс выяснил, что при конъюгации одна клетка является мужским донором, а
другая - женским реципиентом. Клетки-доноры обладают половым фактором F ( от fertility -
плодовитость), который представляет собой замкнутую в кольцо молекулу ДНК. Перенос
генетического материала происходит в одном направлении - от донорской (мужской F
+
) клетки к
реципиентной (женской3 F
-
).
Необходимым условием конъюгации является наличие в клетке-доноре трансмиссивной
плазмиды, продуцирующей половые пили, образующие трубочку, по которой плазмидная ДНК
передается из клетки-донора в клетку-реципиент, в результате чего последняя приобретает
донорские свойства. В случае, когда F-фактор встраивается в хромосому донора и
функционирует в виде единого с ней репликона, то нуклеоид донора приобретает способность
передаваться в клетку-реципиент. Донорские клетки, содержащие встроенный в нуклеоид F-
8

фактор, называются Hfr-клетками ( от английского high frequency of recombination - высокая
частота рекомбинаций).
Процессы генетической рекомбинации у бактерий (трансформация, трансдукция, конъюгация)
различны по форме, но аналогичны по содержанию, т. к. в результате каждого процесса
происходит перенос фрагмента ДНК от одной клетки к другой. При трансформации бактерии-
реципиенту передается свободная ДНК, при трансдукции перенос участка ДНК осуществляется
при помощи бактериофага, а при конъюгации транспортировка участка ДНК происходит через
цитоплазматический мостик между бактериями.
Изменчивость фагов и изменчивость микроорганизмов под влиянием фагов
33333333Фаги, как и микроорганизмы, способны изменять все свои свойства: форму и размеры
негативных колоний, спектр литического действия, способность к адсорбции на микробной
клетке, устойчивость к внешним воздействиям, антигенные свойства. Особенно часто
наблюдаются изменения морфологии негативных колоний, спектра литического действия и
превращение умеренных фагов в вирулентные.
33333333Большие изменения могут наблюдаться в тонкой структуре фаговой частицы — возникают
дефектные частицы, лишенные головки, отростка, нитевидных образований или других
субструктур.
33333333Изменения фагов могут быть наследственными (мутации) и ненаследствеипыми
(фепотипические). Фенотипические изменения зависят от условий, в которых образуются
фаговые частицы. Важное значение имеют изменения, вызываемые клеткой-хозяином, т. е. той
культурой, на которой фаг размножается. Эти изменения большей частью носят
фенотипический характер и касаются преимущественно формы негативных колоний, спектра
литического действия и вирулентности. Под влиянием клетки-хозяина возможны и стойкие
изменения типа мутаций. С помощью разных мутагенных факторов (лучистой энергии,
химических агентов) могут быть получены разнообразные мутанты.
33333333Особый интерес представляют изменения, происходящие при одновременном размножении
на одной и той же культуре двух родственных по антигенным свойствам фагов. При этом в
потомстве возникают частицы каждого из этих фагов и, кроме того, формы, которые приобрели
свойства обоих родителей (гибридные формы).
33333333Под влиянием фагов могут существенно изменяться все свойства микроорганизмов —
морфология клеток, строение колоний, токсичность, подвижность и т. д. Изменения,
вызываемые фагом, могут быть наследственными и ненаследствеипыми. Механизмы,
приводящие к изменению клеток под влиянием фагов, различные. В отдельных случаях фаг
играет лишь роль отбирающего фактора: под его влиянием лизируются все чувствительные к
нему клетки данной популяции и остаются лишь те клетки, которые еще до воздействия фагом
были по разным причинам устойчивы к нему.
33333333При выращивании микроорганизмов в жидкой среде совместно с активным против них
фагом обычно наблюдается следующее. Сначала среда мутнеет, а затем просветляется в
9
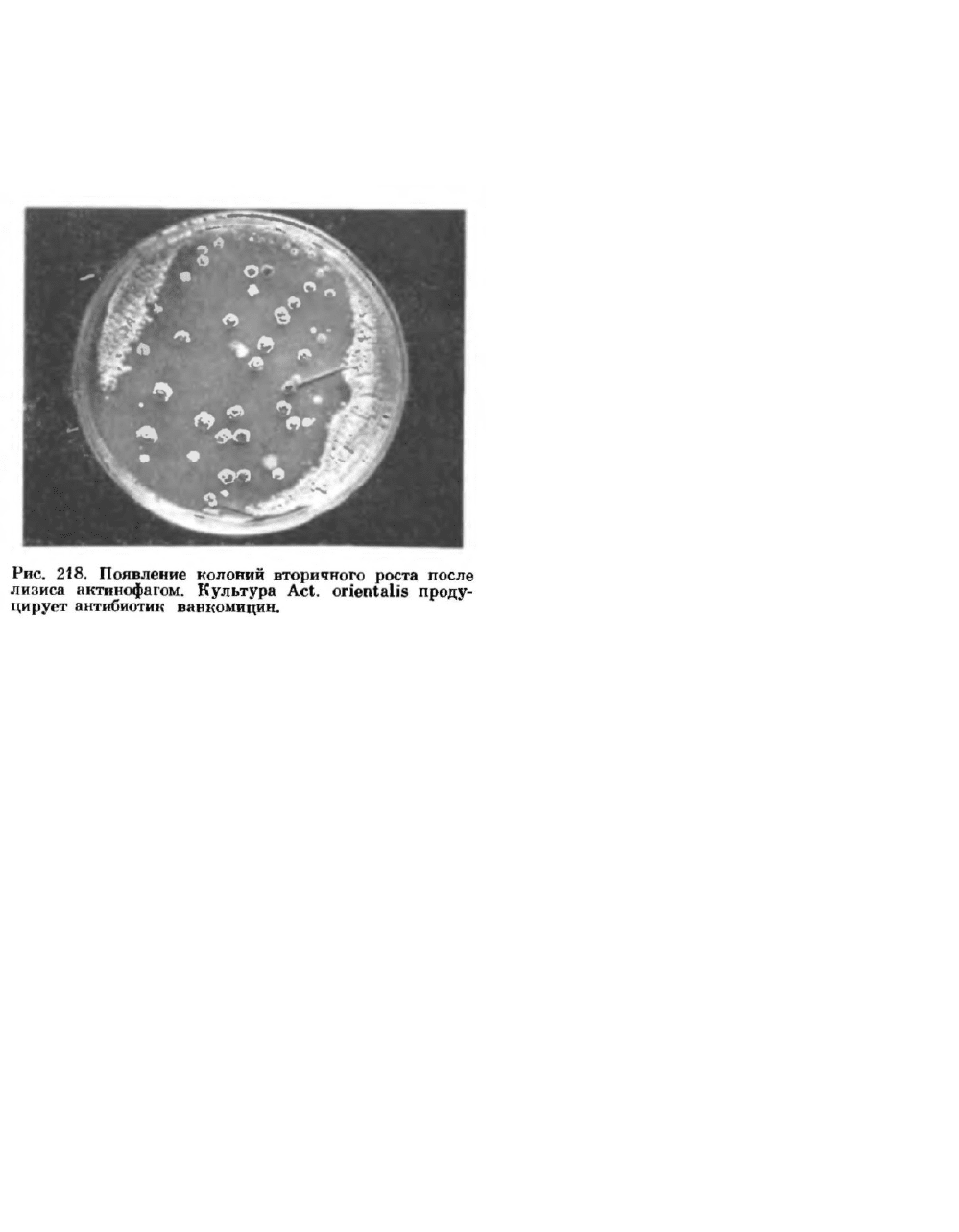
результате лизиса клеток. В ряде случаев через некоторое время (разное для разных микробов и
фагов) культуральпая жидкость снова мутнеет. Помутнение среды происходит вследствие
возобновления роста культуры. На агаризованпых средах при нанесении фага на газон
чувствительной к нему культуры можно наблюдать вначале лизис культуры в местах нанесения
фага, а через некоторое время на лизированных участках появляется рост культуры, обычно в
виде отдельных колоний (рис. 218).
33333333Вырастающие после лизиса микроорганизмы получили название культур вторичного
роста. Анализ культур вторичного роста показывает, что в одних случаях они состоят из
вариантов, ставших стабильно устойчивыми к данному фагу, в других — из нестойких форм.
Культуры, ставшие устойчивыми к фагу, могут одновременно приобрести и ряд новых свойств.
Этим обычно пользуются при получении фагоустойчивых культур для промышленных целей.
33333333В последние годы были выявлены два принципиально различных механизма изменчивости
клеток под влиянием фагов — трансдукция и лизогенные конверсии.
Трансдукция (перенос). При размножении определенных умеренных фагов на
чувствительных к ним культурах фаговая частица захватывает какой-нибудь фрагмент
генетического материала данной клетки. При воздействии этим же фагом на другую
чувствительную к нему культуру он передает новой культуре захваченный фрагмент. Культура,
от которой фаг переносит генетический материал, получила название донора (дающая), а
культура, приобретающая генетический материал,—реципиента (получающая).
33333333При трансдукции фаг играет роль механического переносчика; лизогенизации клетки не
обязательна. Один и тот же фаг может переносить разные свойства.
33333333Трансдукция происходит довольно редко: из одного и более миллионов фаговых частиц
только одна способна осуществлять трансдукцию. При помощи трансдукции удавалось
перенести от клеток-доноров клеткам-реципиентам различные свойства: токсичность,
10