Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.


We shall
not at this
point
deal
in
detail with
the other
functions needed to establish lysogeny,
but we can
just
briefly remark that the infect-
ing
lambda DNA must be inserted into the
bac-
terial
genome (see
Section 19.I7, Specialized
Recombination Involves Specific Sites). The
insertion requires the
product
of
gene
int,which
is expressed
from its
own
promoter
{,
at which
cII also
is necessary. The
sequence of
P, shows
homology
with
P*" in the cII binding site
(although
not in the
-10
region). The functions
necessary for establishing the lysogenic control
circuit are therefore under the same control
as
the
function needed to integrate the
phage
DNA
into the bacterial
genome.
Thus
the establish-
ment of
lysogeny is under a control that ensures
all the necessary events occur with the
same
timing.
Emphasizing
the tricky
quality
of lambda's
intricate cascade. we
now know
that
cII
pro-
motes
lysogeny in another, indirect manner. It
sponsors
transcription from a
promoter
called
Pu,ti-q, which
is located within the
Q
gene.
This
transcript is an antisense version of the
Q
region,
and
it hybridizes with
Q
mRNA to
prevent
trans-
lation of
Q
protein, whose
synthesis is essen-
tial
for lytic development. Thus the same
mechanisms that directly
promote
lysogeny by
causing
transcription of the cI
repressor
gene
also
indirectly help lysogeny by inhibiting the
expression of
cro
(see
above) and
Q,
the regu-
lator
genes
needed for the antagonistic
lytic
nathwav.
The
cro Repressor
Is Needed
for Lytic
Infection
.
Cro
binds to the same operators as
repressor. but
with different affi
nities.
.
When
Cro binds to 003,
it
prevents
RNA
potymerase
from
binding
to P*.,, and blocks
maintenance
of
reDreSs0r.
.
When Cro binds
to other operators at 0* or 0r,
it
prevents
RNA
potymerase
from
expressing
immediate earty
genes,
which
(indirectty)
bl.ocks
reDressor estabtishment.
Lambda
has the altematives of entering
lysogeny
or
starting a
lytic infection. Lysogeny
is initi-
ated by
establishing an autogenous
mainte-
nance circuit that
inhibits the entire lytic cascade
through
applying
pressure
at
two
points.
The
program
for establishing
lysogeny
proceeds
through
some of
the same
events
that
are
required for the
lytic
cascade
(expression
of
delayed early
genes
via
expression
of
N is
needed). We now
face a
problem.
How
does the
phage
enter
the
lytic cycle?
The key
influence
on the
lytic cycle
is the
role of
gene
cro,
which codes
for another
repres-
sor.
Cro is responsible
for
preventing the synthesis
of
the repressor
protein;Ihis
action
shuts off
the
pos-
sibility of establishing
lysogeny.
cro
mutants
usu-
ally establish
lysogeny
rather
than entering
the
lytic
pathway,
because
they
lack the ability
to
switch events
away
from
the expression
of
repressor.
Cro forms
a small
dimer
(the
subunit
is
9 kD) that acts within
the
immunity
region.It
has two effects:
.
It
prevents
the
synthesis
of
repressor
via
the maintenance
circuit;
that
is, it
pre-
vents
transcriPtion
via P**.
.
It also
inhibits
the expression
of early
genes
from
both
Pr and
P*.
This means
that, when
a
phage
enters the
lytic
pathway,
Cro
has
responsibility
both
for
preventing
the
synthesis
of
repressor
and
(sub-
sequently)
for turning
down
the expression
of
the early
genes.
Cro
achieves
its
function
by binding
to
the
same operators
as
(cI)
repressor
protein'
Cro
includes a
region with
the
same
general struc-
ture
as the repressor;
a
helix-2
is offset
at an
angle from
recognition
helix-1.
(The
remain-
der of the structure
is different,
which
demon-
strates
that
the
helix-turn-helix
motif can
operate
within
various contexts.)
As for
repres-
sor. Cro
binds symmetrically
at the
operators.
The sequences
of
Cro and
repressor
in the
helix-turn-helix
region
are
related,
which
explains
their ability
to contact
the
same
DNA
sequences
(see
Figure
I4.2I\.
Cro
makes sim-
ilar contacts
to those
made
by
repressor,
but
binds
to only
one
face of
DNA;
it
lacks the N-
terminal
arms
by
which
repressor
reaches
around to
the other
side.
How can
two
proteins have
the same
sites
of action,
yet
have such
opposite
effects?
The
answer
lies
in the different
affinities
that each
protein
has
for the
individual
binding
sites
within
the operators.
Let us
just
consider
O*,
for which
more
is known,
and where
Cro
exerts
both its effects.
The
series
of events
is illustrated
in
F3fr{Jsf,
x'i.3}.
(Note
that
the
first
two stages
are
identical
to those
of the
lysogenic
circuit
shown
in Figure
14.32.)
The affinity
of
Cro
for
O*3
is
greater
than
its affinity
for O*2
or
O*1.
Thus
it binds
first to
74.20
Ihe cro
Repressor
Is
Needed
for Lytic
Infection
377
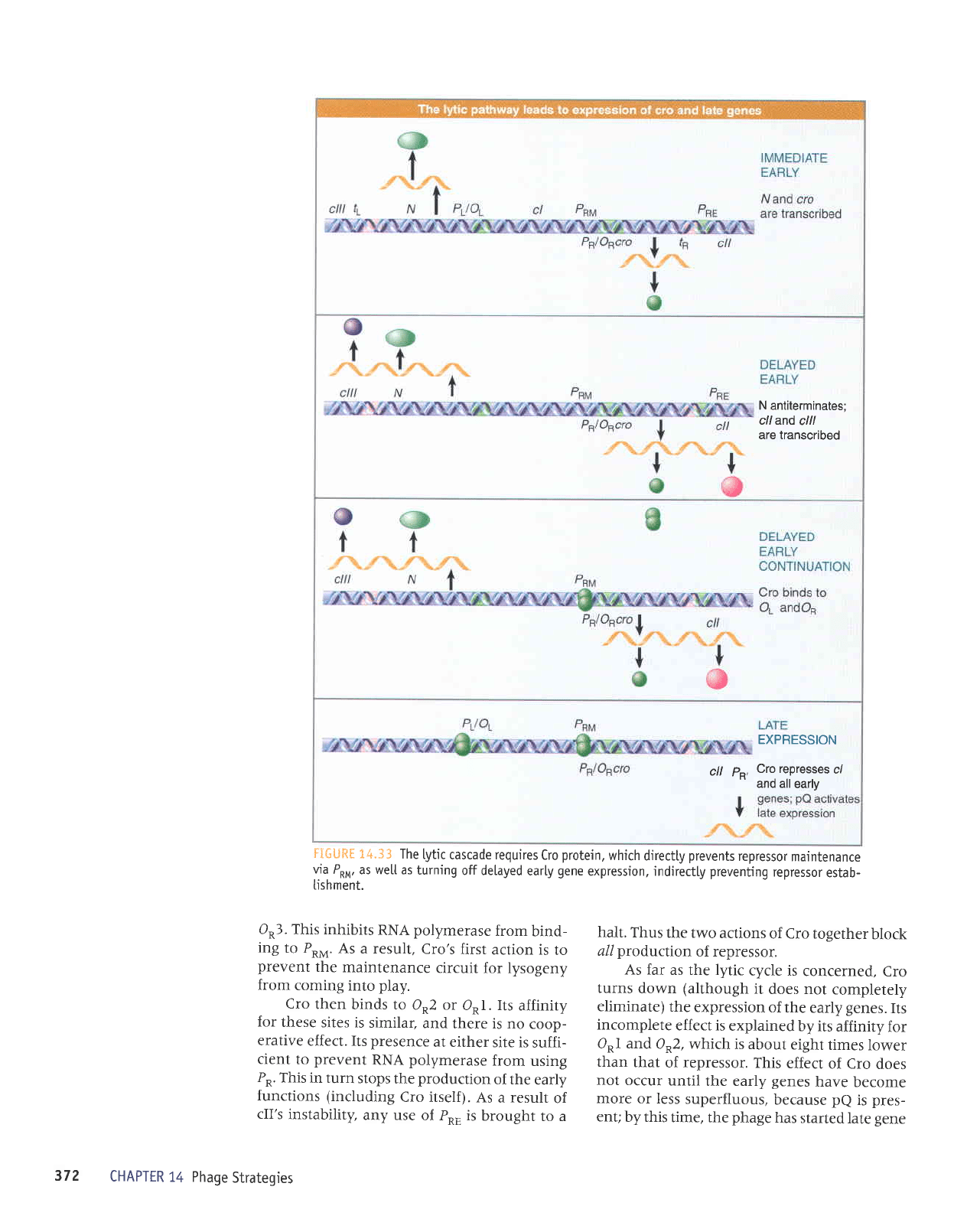
N antiterminates;
cll and
clll
are transcribed
EXPRESSION
cll
Pe,
Cto rePresses
c/
"
and all
earlv
FIGU*E
14.33 The
lytic
cascade requires
Cro
protein,
which
directty
prevents
repressor
maintenance
via
P*r,
as wetl as
turning
off delayed
early
gene
expression, indirectLy
preventing
repressor
estab-
[ishment.
O*3.
This inhibits
RNA
polymerase
from
bind-
ing
to P"*.
As
a result,
Cro's
first
action is
to
prevent
the
maintenance
circuit
for lysogeny
from
coming
into
play.
Cro
then
binds to
O*2 or
O*1. Its
affiniry
for
these
sites is
similar,
and
there is no
coop-
:,H'L'""ff:1il
iliil:ffi?ifJ
n:"J,,
*"
P*.
This
in turn
stops
the
production
of the
early
lunctions (including
Cro itself).
As
a
result
of
cII's
instability,
any use
of
p*,
is
brought
to a
CHAPTER
14 Phage
Strategies
halt. Thus
the two
actions
of Cro together
block
a/i
production
of repressor.
As far
as the lytic
cycle is
concerned,
Cro
turns
down
(although
it
does not
completely
eliminate)
the
expression
of the
early
genes.
Its
incomplete
effect is
explained
by its
affinity
for
O*l and
O*2, which
is about
eight
times
lower
than
that
of repressor.
This
effect
of Cro
does
not
occur
until
the early
genes
have
become
more
or less
superfluous,
because pe
is
pres-
ent;
by this
time, the
phage
has started
late
gene
372

expression and
is
concentrating on the
produc-
tion of
progeny phage particles.
What
Determines
the
Balance Between
Lysogeny and the
Lytic
Cycle?
The delayed early stage
when
both Cro and
repressor are being expressed
is
common to
lysogeny
and the lytic cycte.
The
criticaI
event is whether cII causes sufficient
synthesis
of repressor to overcome the action
of Cro.
The
programs
for the lysogenic and lytic
path-
ways are so intimately
related
that
it is impos-
sible to
predict
the fate of an individual
phage
genome
when
it
enters a
new host bacterium.
Will the
antagonism between repressor and Cro
be resolved by establishing
the autogenous
maintenance circuit shown in Figure
14.32, or
by turning
off repressor synthesis and entering
the late stage of development
shown in
Figure
14.33?
The same
pathway
is followed in both cases
right up to
the brink of decision. Both
involve
the
expression of the
immediate early
genes
and extension
into the delayed early
genes.
The
difference between
them comes down to the
question
of whether repressor or Cro
will obtain
occupancy
of the
two operators.
The early
phase
during which the
decision
is made
is limited in duration in either case. No
matter which
pathway
the
phage follows,
expression
of all early
genes
will be
prevented
as P. and P* are
repressed and, as a consequence
of the disappearance
of cII and cIII,
production
of
repressor via
P*u
will cease.
The
critical
question
comes
down to
whether the
cessation of transcription
from P*.
is followed by activation
of P** and the estab-
lishment
of lysogeny, or
whether P** fails to
become active
and the
pQ
regulator commits
the
phage
to lytic development.
:rJ{rl"iiitrr" :.'rr,
{l
shows the
critical stage, at which
both repres-
sor
and Cro are
being synthesized.
The initial event
in
establishing
lysogeny
is
the binding of
repressor at OL
I
and
Oo I . Bind-
ing at the
first sites is rapidly succeeded
by coop-
erative
binding
of further repressor
dimers at
O12
and O*2.
This shuts off the synthesis
of Cro
and starts
up
the synthesis of repressor
via
Po*.
i
ir:i,ifii
i:;.:r,r The critical
stage
in deciding
between
lysogeny
and
lysis
is when del'ayed
early
genes
are
being
expressed.
If cII causes
sufficient
synthesis
of
repressor,
l"ysogeny
wjttieiutt because
repressor
occupies
the
operators.
Otherwise
Cro occupies
the
operators,
resutting
in
a lytic
cycl.e.
The
initial event
in entering
the
lytic cycle
is
the
binding
of Cro
at
OR3.
This stops
the
lyso-
genic-maintenance
circuit
from starting
up
at
P*r. Cro
must
then
bind
to Ool
or Oo2,
and
to
O.l or Or2,
to
turn
down
early
gene expres-
sion.
By halting
production
of cII
and CIII,
this
action
leads to the
cessation
of
repressor
synthe-
sis
via P*u. The shutoff
of
repressor
establish-
ment occurs
when
the
unstable
cII
and cIII
proteins
decay.
The
critical
influence
over
the
switch
between
lysogeny
and
lysis
is cII.
If cII
is active,
synthesis of
repressor
via the
establishment
pro-
moter is effective,
and
as a
result,
repressor
gains
occupancy
of the
operators.
If
cII
is not active
1.4.21. What
Determines
the
Balance
Between
Lysogeny
and
the
Lytic
cycLe?
373

repressor
establishment
fails,
and
Cro binds
to
the operators.
The
level
of cII
protein
under
any
particu-
lar
set of circumstances
determines
the
outcome
of an infection.
Mutations
that
increase
the sta-
bility
of cII increase
the frequency
of lysogen-
ization.
Such mutations
occur
in c1I itself
or in
other
genes.
The
cause of cIIs
instability
is its sus-
ceptibility
to
degradation
by host
proteases.
Its
level
in the
cell is influenced
by cIII as
well as
by host functions.
The
effect
of the lambda protein
cIII is
sec-
ondary: it
helps to
protect
cII against
degrada-
tion. The presence
of
cIII does
not
guarantee
the
survival
of cII; however,
in the
absence of
cIII, cII is
virtually
always
inactivated.
Host gene products
act on
this
pathway.
Mutations
in the host genes
hflA
and hflB
in
cre a
s e ly s o
geny
-hfl
s tand s f
or high
ft
e
qu
en cy
/ysogenization.
The
mutations
stabilize cII
because
they inactivate
host
protease(s)
that
degrade it.
The
influence
of the
host
cell on the level
of
cII
provides
a route
for the
bacterium
to inter-
fere with
the
decision-taking process.
For
exam-
ple,
host
proteases
that degrade
cII are activated
by
growth
on rich
medium.
Thus
lambda
tends
to lyse
cells
that are
growing
well,
but is more
Iikely
to
enter lysogeny
on
cells that
are starv-
ing
(and
which lack
components
necessary
for
efficienr
lytic
growth
1.
Sum mary
Phages
have
a lytic life
cycle,
in
which infec-
tion
of a host
bacterium
is
followed
by
produc-
tion
of a large
number
of
phage particles,
Iysis
of
the cell,
and release
of the viruses.
Some
phages
also
can
exist in lysogenic
form,
in
which
the
phage
genome
is integrated
into
the
bacte-
rial
chromosome
and is inherited
in
this inert.
latent
form
like
any
other
bacterial
gene.
In
general,
lytic
infection
falls into
three
phases.
In
the first
phase
a small
number
of
phage
genes
are transcribed
by the
host RNA
polymerase.
One or
more
of these
genes
is
a
regulator
that
controls
expression
of the
group
of
genes
expressed
in
the second phase.
The
pattern
is repeated
in the
second
phase,
when
one
or more genes
is
a regulator
needed for
expression
of the
genes
of the
third
phase.
Genes
of
the first
two
phases
code for
enzymes
needed
to reproduce
phage
DNA; genes
of the final
phase
code
for
structural
components
of the
phage
particle.
It
is common
for the
very early
genes
to
be turned
off
during
the later
phases.
CHAPTER
14 Phage
Strategies
In
phage
lambda,
the
genes
are organized
into
groups
whose expression
is
controlled
by
individual
regulatory
events. The
immediate
early
gene
N codes for
an antiterminator
that
allows transcription
of the leftward
and right-
ward
groups
of delayed
early
genes
from
the
early
promoters
PR
and Pr. The
delayed
early
gene
Q
has
a similar antitermination
function
that
allows transcription
of all
late
genes
from
the
promoter
P*,.
The lytic
cycle is
repressed,
and the lysogenic
state maintained,
by
expres-
sion of the cI
gene,
whose
product
is
a repres-
sor
protein
that acts
at the operators
O*
and O,
to
prevent
use
o{ the
promoters
P* and P,
respectively.
A lysogenic phage
genome
expresses
only the
cI
gene
from
its
promoter,
P*no.
Transcription
from this
promoter
involves
positive
autogenous
regulation,
in
which
repres-
sor
bound at
O* activates
RNA
polymerase
at
p
.
RM.
Each
operator
consists of
three binding
sites
for repressor.
Each
site is
palindromic,
consist-
ing
of symmetrical
half-sites.
Repressor
func-
tions
as a dimer.
Each half
binding
site
is
contacted
by a repressor
monomer.
The
N-terminal
domain of repressor
contains
a helix-
turn-helix
motif that
contacts
DNA. Helix-3
is
the recognition
helix
and responsible
for mak-
ing
specific contacts
with
base
pairs
in the
oper-
ator. Helix-2 is
involved
in
positioning
helix-3;
it is also involved
in
contacting
RNA
polymerase
at P*r.
The
C-terminal
domain is
required
for
dimerization.
Induction
is caused
by
cleavage
between
the
N-
and
C-terminal
domains,
which
prevents
the DNA-binding
regions
from
func-
tioning in
dimeric form,
thereby
reducing
their
affinity for
DNA
and making
it impossible
to
maintain lysogeny.
Repressor-operator
binding
is cooperative,
so that
once
one dimer
has
bound
to the first
site, a
second dimer
binds
more
read-
ily to
the adjacent
site.
The helix-turn-helix
motif
is
used
by other
DNA-binding proteins,
including
lambda
Cro.
Lambda
Cro binds
to the
same
operators
but
has
a different
affinity
for
the individual
oper-
ator sites,
which
are determined
by
the
sequence
of helix-3.
Cro
binds individually
ro
operator
sites,
starting
with
O*3, in
a noncooperative
manner.
It is
needed
for
progression
through
the
lytic
cycle. Its
binding
to O*3
first
prevents
synthesis
of repressor
from
P*r,
and
then
its
binding
Io O^2
and
O*I
prevents
continued
expression
of early
genes,
an
effect
also
seen in
its
binding
to O.l
and
Or2.
Establishment
of repressor
synthesis
requires
use of
the
promoter
P*",
which
is
acti-
374

vated
by the
product
of
the c11
gene.
The
prod-
uct of
c111is required to stabilize the cll
prod-
uct against degradation.
By
turning off
cI1 and
c111 expression,
Cro acts to
prevent
lysogeny.
By turning off all transcription except
that of
its own
gene,
repressor acts to
prevent
the
lytic cycle.
The choice between
lysis and
lysogeny depends
on whether repressor or Cro
gains
occupancy of the operators
in a
partic-
ular
infection. The stability of cII
protein
in
the
infected cell
is
a
primarv
determinant of
the outcome.
References
Two Types
of
Regutatory Event Control
the
Lytic Cascade
Review
Greenblatt,
J., Nodwell, J.
R., and Mason, S. W.
(l
99 J). Transcriptional antitermination.
Nature )64,
40),-406.
Lambda
Immediate Earty and Detayed
Earty Genes Are
Needed for Both
Lysogeny and
the Lytic Cycte
Review
Ptashne, M.
(2004).
The Genetic Switch:
Phage
Lambda Revisited. Cold Spring
Harbor, NY: Cold
Spring
Harbor Press
@
Lysogeny Is
Maintained
by
Repressor
Protei
n
Resea
rc h
Pirrotta,
V., Chadwick,
P.,
and
Ptashne, M.
(1970).
Active
form of two coliphage
repressors.
Nature
227, 4l-44.
Ptashne,
M.
(
I 967
)
. Isolation of the
lambda
phage
repressor.
Proc Natl. Acad. Sci USA
57,
306-)l).
Ptashne,
M.
(1967).
Specific binding
of the
lambda
phage
repressor
to lambda DNA.
Nature 214,
2j2-234.
the Immunity
Region
Review
Friedman,
D. I. and Gottesman,
M.
(I982).
Lambda
11. Cambridge,
MA: Cell Press.
@
The DNA-Binding
Form of Repressor
Is a
Dimer
Researc
h
Pabo, C. O. and
Lewis,
M
(1982).
The operator-
binding
domain of
lambda repressor:
structure
and
DNA recognition.
Nature
298, 44)-447.
The Repressor and
Its 0perators
Define
Review
Repressor Uses
a
Helix-Turn-Hetix
Motif
to Bind
DNA
Resea
rch
Sauer,
R. T. et al.
(
1982
)
. Homology
among
DNA-
binding
proteins suggests
use
of a
conserved
super-secondary
structure.
N ature
29 8,
447
45
1 .
The Recognition
Hetix
Determines
Specificity
for DNA
Resea
rch
Brennan,
R. G et
al.
(
I 990).
Protein-DNA
confor-
mational
changes
in the crystal
structure
of a
lambda Cro-operator
complex.
Proc
Natl'
Acad.
Sci USA 87.
8165-8169.
Wharton.
R. L
,
Brown,
E.
L., and
Ptashne,
M.
(1984).
Substituting
an cx-helix
switches
the
sequence
specific
DNA
interactions
oI a
repressor. Cell
)8,
361-)69.
Repressor
Dimers
Bind
Cooperativety
to the 0perator
Resea
rch
Bell,
C.
E., Frescura,
P., Hochschild,
A.,
and
Lewis,
M.
(2000).
Crystal
structure
of
the
lambda
repressor
C-terminal
domain
provides
a model
for cooperative
operator
binding.
Cell
I0l, 801-81
I.
Johnson,
A.
D., Meyer,
B. J.,
and
Ptashne,
M.
(197
9). Interactions
between
DNA-bound
repressors
govern regulation
by
the
phage
lambda
repressor.
Proc. Natl.
Acad. Sci USA
76'
5061-5065.
Repressor
at 0*2
Interacts
with
RNA
Potymerase
at
P*t
Resea
rc h
Hochschild,
A.,
Irwin,
N.,
and
Ptashne,
M.
(1983).
Repressor structure
and
the
mechanism
of
positive
control.
cell
J2,
319-325.
Li, M., Moyle,
H, and
Susskind,
M' M.
(1994). Tar-
get
of the
transcriptional
activation
function
of
phage
lambda
cI
protein. Science
26),75-77.
Cooperative
Interactions
Increase
the
Sensitivity
of
Regutation
Ptashne,
M.
(2004). The Genetic
Switch:
Phage
Lambda
Revisited.
Cold
Spring
Harbor,
NY:
Cold
Spring
Harbor
Press.
Resea
rch
Bell, C. E. and
Lewis,
M.
(2001). Crystal
structure
of the
lambda
repressor
C-terminal
domain
octamer.
J Mol.
Biol.3l4,
1127-1136.
Dodd, I. B.,
Perkins,
A. J.,
Tsemitsidis,
D., and
Egan,
J.
B.
(200I).
Octamerization
of
lambda
CI repressor
is needed
lor effective
repression
of P(RM)
and
efficient
switching
from
lysogeny.
Genes
Dev.
| 5,
)Ol3-3022'
References
375

The
Repticon
CHAPTER
OUTLINE
1
l
Introduction
3E7.,
Repticons
Can Be
Linear
or Circutar
r
A replicated
region
appears
as an
eye within nonreplicated
DNA.
r
A replication
fork
is initjated
at the
origin
and then moves
sequentiatly
atong
DNA.
.
Replication
is
unidjrectional
when
a singte reptication
fork
is
created
at an origin.
.
Reptication
is
bidirectjonal
when
an origin
creates
two
repti-
cation forks
that move
in
opposite
directjons.
Origins
Can Be
Mapped
by Autoradiography
and Etectrophoresis
.
Reptication
fork
movement
can
be detected
by
autoradiogra-
phy
using radioactive
putses.
.
Replication
forks
create Y-shaped
structures
that
change
the
electrophoretic
migration
of DNA fragments.
Does
Methylation
at
the
Origin Regulate
Initiation?
o
onC contains
r'
carr
rteven
;iA;
repeats
that
are
methylated
on
adenine
on
both
strands.
r
Replication generates
hemimethylated
DNA, which
cannot
initiate
reotication.
Replication
0rigins Can Be
Isolated in
Yeast
.
Origins
in S.
cerevisiae are
short A-T-rich
sequences
that
have
an essentia[
11-bp
sequence.
e
The
ORC is
a comptex
of six
proteins
that
binds to
an ARS.
Licensing
Factor
Controts
Eukaryotic
Rereptication
r
Licensing factor
is necessary
for
initiation
of reptication
at
each origin.
r
It is
present
in
the nucteus
prior
to reptication.
but is
inac-
tivated or
destroyed
by
replication.
o
Initiation
of another
reptication
cycle
becomes
possibte
onty after
licensing factor
reenters
the nucteus
after
mitosis.
Licensing
Factor
Consists
of MCM Proteins
.
The
ORC is
a
protein
complex
that is associated
with
yeast
origins
throughout
the cetl
cycle.
e
Cdc6
protein
is
an unstabte
protein
that is
synthesized
on[y
in G1.
e
Cdc6
binds
to ORC and
a[tows MCM
proteins
to
bind.
o
When reptication
is initiated.
Cdc6 and MCM
proteins
are
disptaced. The
degradation
of Cdc6
prevents
reinitiation.
o
Some MCM
proteins
are in
the nucteus
throughout
the cycte,
but others
may
enter onty
after mitosis.
D Loops
Maintain
Mitochondria[
Origins
o
Mitochondria
use
different
origin sequences
to initiate
reptication
of each DNA
strand.
o
Reptication
ofthe
H strand is
initiated
in
a D [oop.
r
Replication
of the L
strand is initjated
when its
origin
is
exposed
by the movement
of the first reptication
fork.
Summary
.
There is
a 13-minute
detav before
the
remethvtated.
fflf
,eneats
a'e
0rigins May
Be
Sequestered
after
Reptication
.
SeqA
binds
to hemimethylated
DNA
and is
required
for
detaying
rerep[ication.
.
SeqA may
interact
with DnaA.
r
As
the
origins
are
hemimethytated
they bind
to the
cetl
membrane
and
may
be
unavaitable
to methytases.
o
The
nature
of the
connection
between
the
origin and
the
membrane
is
sti[[
unctear.
Each
Eukaryotic
Chromosome
Contains
Many
Repticons
.
Eukaryotic
replicons
are 40
to 100 kb
in
tength.
e
A chromosome
js
divided
into
many repticons.
o
IndividuaI
repticons
are
activated
at
characteristic
times
during
5
phase.
o
RegionaI
activation
patterns
suggest
that repticons
near
one
another
are activated
at
the same
time.
376

LLE
uoqlnpollul
I'EI
oc
zluelrs
sarurleuros surSuo eruos eJe Jo
'pesn
s^eule sur8rJo
IIe
JrV
'alr^J
IIJJ
z(rarra ur
Jrues Jql sr uorlerrldar
Jo
uralled Jqt JeqlJqM
,lt.ou>l
tou
op a14
'suorrldar
uaa.lrtaq drqs
-uorlelar
eql
tnoqe
uorteurroJur Jlllll a^eq
IIIIS
aM
1nq
'suorlldar
lenprlrpur
Jo
uorlrnrlsuof, Jqt
lnoqe
uoueuJolur
DJIIo)
or
un8aq J^eq
JM
'patalduror
uaaq seq suorrldar
11e
Surlerrldar
yo
ssarord
JJrlue aql uaqm
aleJrpur
ol
lsrxa lsnru
leu8rs
raqtoue
os
,{ltuapuadapur
palenltf,e
JJe
suoJ
-1dar
LuepJurt
puo)as
p
eJrI
lou
op suortldar
teql
JrnsuJ o1 suorrldar
palelldaruou
ruoJJ
palerrldar qsrn8urtsrp
lsnru
leu8rs
aruog
'a1tft
yac
4)0a
w iluo ueLli alow
0u
pawru)a
aq
lsnut
LpDa
puv
'pouad
paDelord
.,i.prey
e rJAo
petenrlle
eq
lsnru
uorrldar
qreg
'.{.lsnoauellnrurs
Jlrlf,e
lalJ,vloq
'lou
are
Laql
'apAr
1ar
auo Surrnp
perrJ
aq
lsntu
euosoruoJqf,
p
uo suoJrldar aqt
Jo IIV
:loJtuoJ
Jo
rual
-qord
aql
ot uorsuJrurp
raqloue
sppp srr{J
'uorl
-elldar
Jo
slrun.dueru sapnpur uorle8ar8as
;o
lrun
Jql snqt
lsuorrldJJ
Io
rJqrunu a8rel e suret
-uoJ
Jurosotuorq;
rrlor(re>lnJ
qlpg'uorlerrldar
Jreql ur uJJs sr sauoua8 rrlo.,i:e1na
pue
IprJJpeq
Jo
uorlezrue8ro
aqt ur eruerelJrp roferu
y
ilil
a41 ut
flsnowouolna
palat1dat
aq un ut6uo ua suruiun
io4i
alwapw
VNe
[uV
:uorlrurJep
eql JSJaAeJ ol sr
'JJoJaJeql
'uor[dar
llo.{re>lo:d
Jql MarA o1
,{em
Jellaq e sdeq:ad
'ap.dr
snorlrJJur
ue Surrnp sarurt
duBru
alellpr
ot JIqp sr
'snql 'pue
uorlldal e sJlnqtsuo) osle
VN(
snrrl ro
a8eqd
qrpg
'JruosorxoJqJ
lerrJl
-Jeq
aqt ueql
sardor
Jo
Jaqunu ralear8 e ur
tue
-sard
sr
tl
uar{M
'IorluoJ
Ldor;l1nur rJpun Jq
Leu
1r
ro
'salerrldar
JruosoruoJqJ
IerrJDpq
Jql
arurl drazra
aruo salerrldar
11
1pql
sueJru
qJlqM
'lorluor
Ldor a13urs ,uoqs ,{eru uorrldar
pru
-se1d
y
'(7'71
arnSrg aasl uotqdat
awndas
a
safi1
-IISUU
1ot41
awOuaq
y51q
JalruJD sn7ut7u7lna ua sr
ptwsold
y'spruseld
Jo
ruroJ
eql ur uorteturolur
rrlaua8
Ieuorlrppe
ureluoJ .4.eu erralreg
'z(dor
a13urs
pelleJ
sr
IorluoJ
uorlerrldar
Jo
adzlt srql os
'Jruosoruorqr
a13urs e seq urnrJel
-req
proldeq
qJpA
'uorsrlrp
11ar
^drana
JoJ eJuo
'auoua8
JrrluJ Jql
Jo
uorletrldar srosuods ur8
-rro
a13urs
e
le
uorlerlrul
'aprJuroJ
uorle8ar8as
pue
uoner{deJ
Jo
stun Jql snql
luorqdar
a18urs e
selnlllsuof,
IIar
rUo.,hB>{ord e ut
auouaS
y
('areqMesle
pepoJ
aq Leu
(s)urarord
rotelnSar ?u:gte-suaq lur8
-uo
ue suleluo)
leql
uorlerqdJr
Jo
lrun
p
JqrDSep
ot sJurosoruorqr
rrloLrp>lnJ
ol
parldde
,(11e
-nsn
sr
,,uorrldar,,
'Jale,lror{ 'MoN
'uraloJd
Jolpln
-3ar
aqt ro; Surpor aua8
aql
pua
ur8rro aq1
qloq
Surssassod
tlun
e se
tl
pJMarA
[sato.drelord
ur]
uorrldar er{r
Jo
uortplnruJoJ pur8rro
aq1)
'sJprsar
1l
qrlqM
uo
vN(I
Jo
JInrJIoru
ieqr
'(po
DeJJp
01 elqe
'etIS
Suttre-sr
e sr ut8rro JqJ
'uo)
-rtdar
leqt;o
ged
se
palerrldar
sr-snururJet
p
dq
ur8r.ro
ue uorJ
paleredas
1ou
Llasoard a.roru
'ro-ur8uo
up ol
pJqJelle
aruanbas .d.uy
'sdots
uorterqdar
WIqM
tp
snurruJal
e eneq osp ^,{eur
11
'pJlprlrur
sr uorlerlldal
qJrqm
1e
u161ro
ue
seq
tI
'uorle)
-qdar
roy
pepeeu
stuJrxele
IorluoJ
eqt
Jo
uors
-sassod
s1r Lq
pauryap
sr uorlldar aq1
'a1rub
1ar
qrea
ur
'aruo
.d1uo
pue
'aJuo
,,seJrJ,,
uorqda:
qreg'uorqdal
eqt
pJIIer
sr srn)Jo uoperqdar;o
De
Ipnpr^rpu
ue
qlrqM
uI
YNo
Jo
uun
JqJ
lpapu8rs
uorleurrrrJet sU sr
,r,roq
pue
ssarSord
stl slorluol
teq6
Zpatpltlul
uoperlldar;o ap,{r e
sr MoH
'uorlerqdar
VNq1
Jo
uorleln8ar eql
qlrM
pJurJJuoJ
JJp JM
'retdeq:
srql ul
'srsolnu
lp
IIJJ
eqt
Jo
uorlezrue8roar aql z(q
paqstdurone
sr uorsr^rp
Jo
lJe
eqJ
'slue^a
uollPrtlur
IPn
-pplpq
^dueru sanlonur
qJrqM
pue
'sJnJJo
srs
-aqtuz(s
VNq1
qrlqM
Surrnp
porrad paperlord
e
'aseqd
S
Jo
upts
aqr,{q
pagrrurpr
sr uorte)r1dar;o
uorterlrur
's1ar
rrloLre>lne uI
'o,tal
olur
IIaJ
Jql
sephrp
pup
IIeM IIJ)
eqt ruorJ s.uor8
leqt
runtdas
e
yo
tuarudolJlJp
aql Lq
paqsqdruoJJe
sr uors
-IAIp
Io
ssaro.rd aqt
pue
'Jurosoruorq)
IerJJl)eq
Jql uo alrs anbrun e 3ut,L1o,rur
lua.aa
a13uts
e
sr
uorlelldar
Jo
uorlerlrur aql
'salorfue4ord
u1
'Jruosotu
-orqJ
Jql sr uorle8ar8as
Jo
lrun
aqJ
'11ar
ralqSnep
qJea
ot euo
petp8arSas
ueql eJe saruoua8 aterlldnp
eqJ'uotsr^rp
IIel
JoJ ra33rr1 e aprnord
,{eru
uorler
-qda.r
yo
uo4aldruor aql
'paJpul'pa1a1d
-uro)
uJJq spq
IuJAJ
uorlerrlda.r Jqt
lrlun
rn)Jo ol
pallnurad
Jq
louupr
uorsrlrp
luanbasuor
Jql
'spJJJord
uorlerlldar
;1
.
'pala)tlanp
uaaq saLl aw0ua6 anlua a4l
plun
sanuuun
u
'paqa$
satl uo4atqdal nuo
'uorteltlul
Jo
a8els aql
le
palloJluoJ
sr uouer{dag
'uor1
-elldar
VNC
Jlelltur
ot
tou
ro JJqlJqM
uo suorsrJJp
Jo
sJrJJs e
{q
paunu.ralap
sr
sateraua8
IIJJ
e
teqt
sluepuJJsJp
Jo
JJq
-rrrnu
Jq1
'lurodpuels
srql ruoJ{
'ultstrtp
ntlunt
a ol
(t110{w4na
n t4ofua4otd)
17ac
aLli sllwutn uo4at1dat
VNA
t0
uoqvqtul o
:JIJAJ
IIJJ
eql
Jo
uonrpuoJ Jr{l
qll.,rt
uorteJrldar
yo
alels eql
aredruor ol
pJSn
are saldrourrd
praua8
o,n41
;apLr
IIaJ
aqt ot
pJ>lurl
uorlerrldar
Jo
pe
eql sI
zlroH
'uorsrlrp
lar
fuana JoJ Jf,uo ,{.1asoard
paler
-qdar
aq
tsnru
ouroua8 arrlua aqt
'(sato,{re1na
ur se) saruosouorr{J Lueru seq ro
(sa1ol.re1ord
ur se) eruosouroJqJ auo
r(1uo
seq
IIeJ
e reqlaq6
uoqlnporlul

uorrloeu
eql
9I
ulldvHl
gLE
'ur6uo
aql
lP
pauloJ
olP
sl.loJ
uor+Prrlda.l oMl to
auo loqloqM
uo ourpuodap
,lpuorl
-lerrprq
io
lpuorJrerrpr.un
aq /\eu
suolrtdeu
*''jl-
3i*gllii:j
Jqt
'uorlpJlldar
puorpaJrprun
,{q pareraua8
I
'sarnlJnrls
oMl
Jo
JJqlrJ
luasa,rdar
uel a,{a :ql
'i:
ti
F:.iiii:iij
ur
paDrdap
sV'uortellder
leuortf,eJ
-Iplq
puP
IeuoIlJJJIprun
uJaMlsq qsmSuusrp
10u
saop
aLa uorlerqdar
p
Jo
JJueJpedde
aq1
.SUOIlJEIIP
JUSoddo ut ut8tro
Jql ruorJ .,(e.nae paatord
r(aqr
lperuJoJ
are
s>lJoJ uorlelldar
o^{t
'uolleJIIdJJ
IeuolDeJrprq
uI
'vNC
er{l Suole
spaarord
pue
ur8rro eql sa^pJl
>lJoJ
uorleJrldar
auo
'uorleJ
-r1dar
leuogreJrprun
u1
'ur8uo
eqt
uorJ
tno
las
s>lroJ uorleJrldar
o..tal ro
auo JJqleqM.dq
pau[u
-ralJp
sr
lue^e;o
adLl
JqI'uolle)qdar puol1
-Jarlprq
ro
uolterlldar
lpuollJeJrprun
rJqtra
uels
ot
pasn
aq,{.eru u€rro
aq1
'ut6uo
a417a
rutod
Cuunis s11 wotl
y111q
aLlj 6u0lu
{11a4uanbas
satow
4totuo4atqda;
y'(1urod
8qmo,r3
Jql sp uMou>I
osle seullJluos)
>1,roy
uogler;1dar
Jql
pelleJ
sr srnrJo
uolterrldar
q)lqM
le tulod
aqJ
'pJruJoJ
aneq saxaldnp
ratq8nep
oM1 Jqt JJeqM
uor8ar
palerrldar
eql otur
suado srqt
lxaldnp
IeluJrpd
Jql
Io
slsrsuor
uo6ar
palerlldaruou
aq1
'YNO
pJlel[daruou
Jr{] ur{lr.&\
a1[a
uol1er11dar
e se sreadde
uor8ar
palerrldar
aql .{dorsontu
uollJale ,{.q pa,nat,t
sl
VNC
Surlerrldar
uaqM
]Pql
sMoqs
:.":':!.
:ili+:jiri
'suorSar
Jo
sad.dt o.ta.l
seq uorlerr1dar ur
pa8e8ue
VNO
Jo
JIn)JIoru
V
a^e uoLlerqder
p
sp uaes
"
Yil8
5:];lilj:i'?fl.|;]t1:,]']
'su0qlarrp
alrsoddo ur o^oru
leql
syoJ uotlerqder oltl
saleen urbuo up uaqM
leuoqlellprq
st uotlertldag
r
'urbuo
ue
lp
palporl
sr
yo;
uoLlerqder
e16urs e uaqM
lpuolparrplun
sr uorlerqdag
o
'y16
6uo1e fllequanbas
se^ou uoql
pue
urbuo
oql
le
palprlrul
sl
l.lo1
uotlerqder
y
o
'vNc
palelrl0alu0u
urqlwr eAe ue sp sreadde uoLbar
palerqdar
y
.
JPlnlrrl
ro
leoull
aB uPl
suollldau
'errel)eq
ralq8nep ot
sJruoue8 eql
Sulle8aJSes
ro;
elqrsuodsar sr
teqM
pue
UoISIAIp
IpI
-JalJpq
qu1!t
pJleulpJooJ
sr JruoueS eql
Jo
uop
-erqdar
Moq
Jo
uorlsJnb eql
ol uJnl a.,n
'ur8rro
Jqt
Jo
uonerJprsuoJ aqt
3uu,ro11oy
:a1e1 [aqi
teqt
sruJoJ snorJe^ Jql
pue
suorlldar
Jo
uolllnJlsuol
rrseq Jql Surraprsuor
dq
uels
a6
'uotlerqdar
ro; snleredde Jqt
Jo
surato.rd
aleudordde
aql
^dq
pazruSorar
are ,{aql
,lroq aulruJelep ot
pup
surSrro sp uorlJunJ
]eql
saJuanbas
aq1 eurJep
ot sr uortsanb ,{a1 aqr
'sruats^s
JSJqI
IIe
uI
'elJAJ
IIel
aqt
01
pJleler
Jq
tsntu
uorlerrldar
vNe
elleue8ro
1o
loJluo)
eqt
pue
'11ar
rad
ylqq
a11aue3Jo
qf,eJ
;o
satdot
a1dr11nru JJe arJqI
'urnrrJlJeq
rad sardor
a1d1l
-lnru
ur
lsrxJ teqt
sptuseld
J>III aJou
palep8ar
aq,{eru stseldorolqr
pup
plJpuoqJolllrr
Jo
VN(
eqt
'lortuoJ
;o
adLl
.,(dor-a13uts
p
JAeq
qJIqM
'sJruosourorql
JeJIJnu
qll.&r
ISPJIUOI
uI
zluaql
saqsm3uqsp
leq,n
'sur8uo
Jo
sJssep
tuereJJlp
JJe
ereql
II
ZrepJo
Jrups
eql uI
erIJ s,{.elt1e sur8rro
Nt0rHo
NO|IVCt-ld3H rVNOtIClHtOtE
NOI-LVCt'td3
u rvNo
tl-cf H to I
Nn
Nr9rHO
VNO
VNC
peiecrlderuoll
a;ta
uo$eslldaH
pelectldetuoll
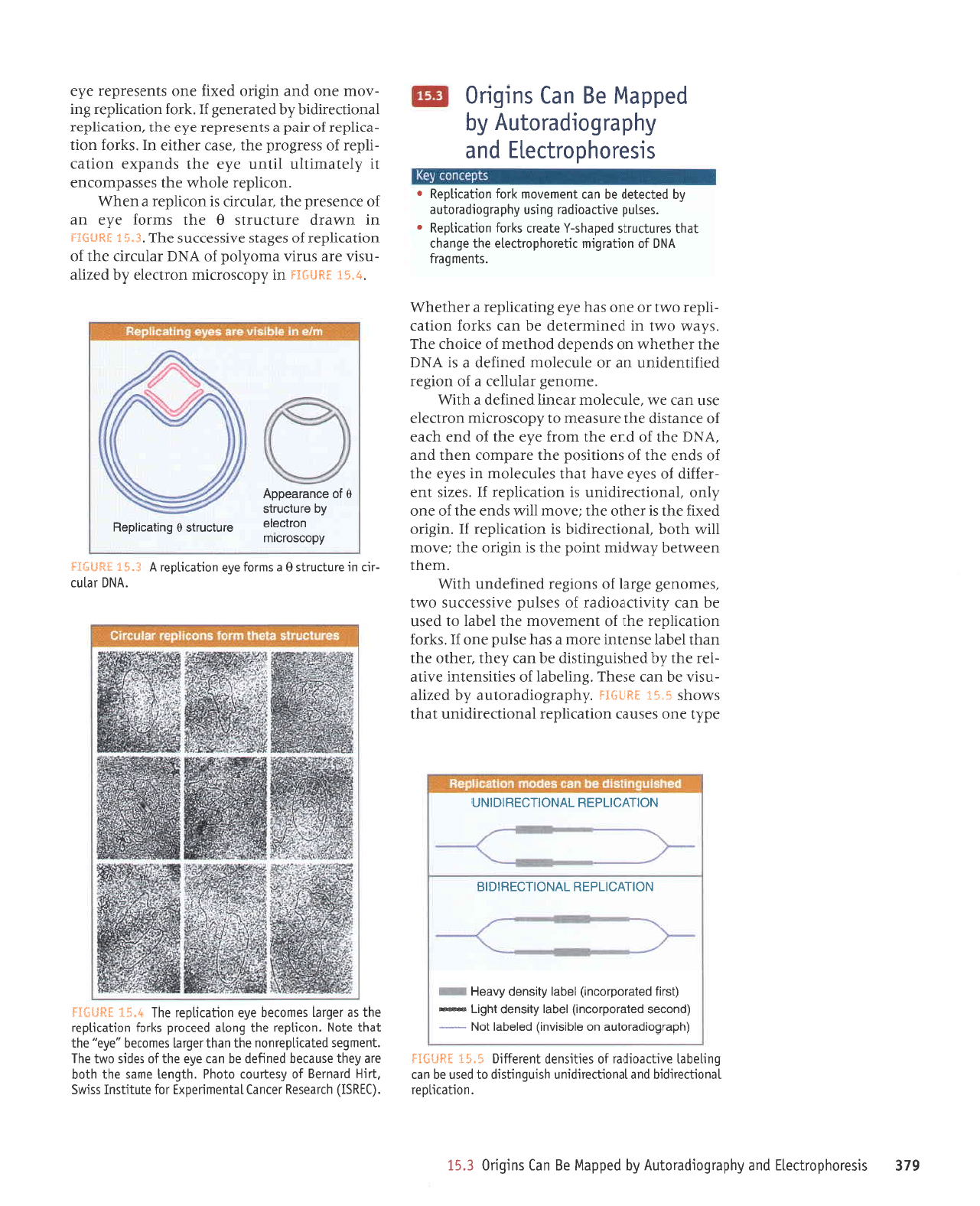
6Lt
srsaroqdorllall
pup
^qdprborpprolnv r{q
padde6l
ag uel surbug
g'E1
'u0qeltl0el
leuoq)olrprq
pup
leuoqlarrprun
qsLnbuqsrp
01
pesn
eq uel
6uqaqel o^rlleorppr
Jo
sarlrsuap
luoro#r6
g'Ei
3'dn$g
(qder6orperolne
uo e;qrsrnur)
palaqel
loN
-
(puoces
palerodrocur)
laqel
Iltsuep
lq6t1
*
(1sr11
palerodrocur)
1eqel
[lsuep {nea;1
NOrtvcndf u lvNotlcf
HrorS
NOtrvct-td3u]vNoU-c3uto tNn
adAl auo sesneJ uorleJrldJJ
lpuorlf,Jrrprun
leql
smoqs
*"Ei
:EilSIj
',{qderSorpe:orne
,{.q
przue
-nsrl
Jq uer JseqJ
'3u11aqe1
Jo
sartrsuJlur enrle
-ler
eqt.{.q
paqsrn8ultsrp
eq uer Laqt
'reqlo
aql
ueqt
IJqBI
JSuJtul JJoru
p
seq aslnd Juo
JI's>lJoJ
uorlBrlldat Jql
Jo
tuJrua^oru
3q1
Iaqel
o1
pasn
Jq ueJ ,{.lrnrlreorper
;o
saslnd enrssaJ)ns oul
'setuoua8
a8rel
yo
suor8ar
pJurJJpun
qu6
.luaql
uJeMlJq ^delrpnu
lurod
aqt sr urSrro aql
ie^ou
IIrM
qtoq
'leuorl)Jrrprq
sr uouerlldar;1
'ut3rro
paxrJ
Jql sr Jeqlo er{l
la^oru
IIrM
spuJ eql
Jo
Juo
.d1uo
'leuorl.lJJrprun
sr uorlerrldal
JI
'sJzrs
luJ
-rJlJIp
;o
sar(a eAPq
leql
selnJJloru ur sa,{.a aql
Jo
spue aql
Jo
suortrsod aqt aredruor uJqt
pue
'vNQ
eql
Jo
pur
rql ruorJ a,,{a
aqt
Io
pur qJPe
Jo
Jf,uelsrp
aql JrnseJru o1
^ddorsonrru
uoJlJela
Jsn ueJ J,/!t
'JIn)JIour
rpeurl
paurlep
p
qllM
'eruoua8
relnllJJ
e
yo
uodar
perJrluJprun
ue ro elnJelotu
peurJep
e sl
yNQ
Jql raqlaqM
uo spuadap
poqtJur
Jo
JlroqJ JqJ
'szi.eazr
oazr.l
ur
peururrelep
eq ueJ s>lroJ uorleJ
-r1dar
o,u1 Jo Juo seq al.a Surlerqdar
p
JeqtJqM
's1uaube4
y116
;o
uoqerbru rrlaroqdorlrala
aq1 abueqr
lPql
sernltn.lls
paopqs-A
alpall slloj uogeltloel
r
'saslnd
anrlceorper 6ursn Aqder6orperolne
r{q
papalap
aq upl
luauo^or!
lto1
uoqerqdaX
r
srsajoqdorllall
puP
fiqder6orpProlnv
Aq
'(flUSI)
ql.lpasau
rolupl
lpluauuedx3
ro; alnltlsul sst/v\S
'lrrH
pleurag
1o
Aselnor o1oq6
'q16ua1
aups aql
qloq
are
r\eq1
asnpraq
peulJap
eq uer afia aqlJo sapts oMl aql
'lueubas
palplrldaruou
eq1 ueql rebrel souolaq
,,ar{e,,
aq1
lpql
aloN
'uocLldar
eq1
6uo1e
paarord
s1to1 uoLlerqdar
aq1 se tebrel sauotaq aAa
uoqetqder eq1
+.8
[
]Hfi*:l
'vN0
lPlnl
-lo
ur arnllnlls
e
p
suuo1
aAa uo4erqdar
V €'$t IHfi+ij
{docso.rctu
uorlcele
r{q ernlcnrls
g;o
acuereeddy
oJnlcnrls
6
6urtecrldeg
'?'S{
}Un1ru
ur
,{.dorsonrru
uoJDele Lq
pazle
-nsrl
are snrrn
eruoLlod
yo
y1qq
relnJJrJ
JrIl
Jo
uouerqdar
Jo
saSels
JAISSaJTns
aqI'€"9
il
SSnSIj
ur uueJp aJnlJnJls
e
Jql sruJoJ J^J up
;o
aruasard eq1
'relnJJD
sr uolldar
p
uJqM
'uorqdar
eloqM aqt
sJSSpdtuoJuJ
1r
[1a1erur11n
Irlun
aii.a aql spuedxa
uorle)
-qdar
;o
ssarSord Jqt
'Jsef,
JJqtlJ
uI
's>lJoJ
uorl
-errldar
yo
rrcd e sluasardar ale aql
'uorlecrldar
IeuorlJJJrprq
r{q
palerauaS
JI
'{roJ
uoperqdar 3ur
-Aoru
euo
pue
ur8rro
pJXrJ
euo
sluasardar a,{a
padde6
ag uel sur6rr0
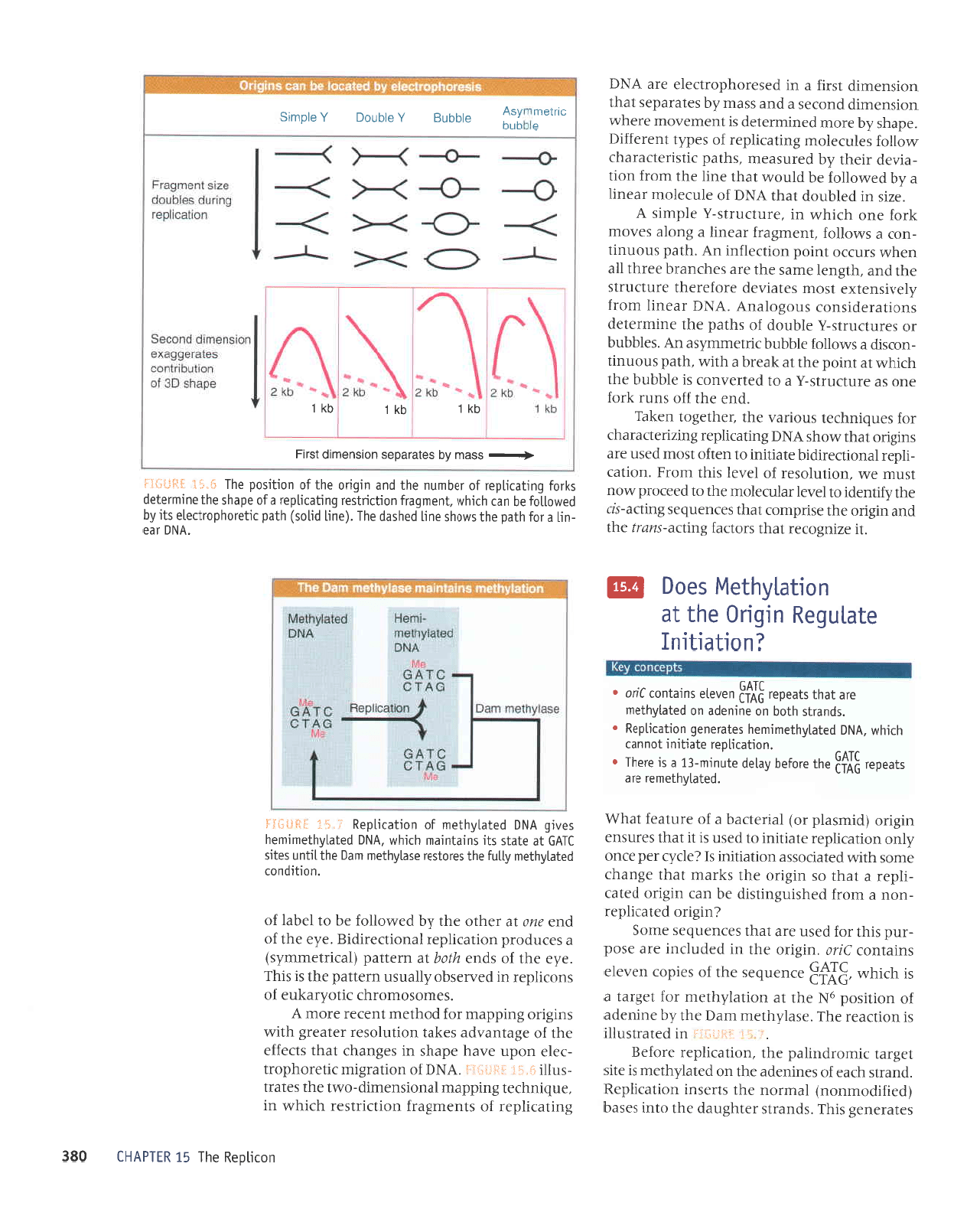
seleJJuJS
srqJ
'spueJls
talqSnep
Jql olur
sJSpq
(pJrJrpouruou)
Ieurou
Jqt suJsur
uorlpJrldJg
'puPns
qJeJ
Jo
sauruepp
Jr{] uo
palelr{qtau
sr alrs
la8ret
rtuorpu[pd
Jqt
'uorle]rldar
aroyag
:
:.i
::!ill:i-t
UI
pJleJlsnlll
sr uortJeJJ
aq1
'ase1.{qtJu
rupc
Jq1 ^q
Juruepe
yo
uorlrsod
qN
aql
le
uortelz{qtJlu
roJ
1a3re1
e
r\vTa
sr
qrrqM
'5jVrd
a:uanbas
aqt;o
sardor ur^ele
sureluoJ
3rto
'urSrto
Jql
ur
pJpnl)ur
aJe
esod
-rnd
s1q1 JoJ
pesn
Jrp
teqt
seJuJnbJS
aruos
2ur8rro
paterrldar
-uou
e
uoJJ
pJqsrn8urlsp
Jq uel ur8rro pater
-r1dar
e
lpqt
os ur8rro
aqt
s>llptu
lpql
e8upqJ
Jruos
qlrM pJlprf,osse
uorlerlrur
s1
;a1t,{r
rad
aruo
^dpo
uoqerlldar
alerlrur
01
pJSn
sr
lr
teql
sJJnsuJ
ur8rro
(ptuseld
ro)
IeueDpq
e
Jo
rrntprJ
teq6
slea
d a.r
?Y.11
a
qr
ar
ole
q
rel
ap rrr .|,tJ:]iultT
:|tfi
.
llv9
'uorlPlrloal
aleqrur
louuel
qllqM'VN0
palelfqlaLururaq
seletaueb
uorlerqdag
.
'spuptls
qloq
uo ouruopp
uo
pa1e1[q1aLu
a.re
teql
sleader
lf'!
uenate
sureJuo)
JUo
o
auoqeqrul
atPlnbau
ur6u0
aql
le
uoqPl^qlal^,
seoo
@
'1r
azruSorar
leql
sJopeJ
7ur1te-sua4
aql
pue
ur8uo
aql asrrduor
leql
saruanbas
SurDe-sr
Jqt ^JrtuJpr
01
IaAJI
JelnJelolu
aqt ol
paarord,vrou
lsnur
JAt
'uorlnloseJ
Jo IJAJI
Srql
luOJC
.uorleJ
-ITdJr
IeuorlJJJrprq
elerlrur
ol uauo
lsotu
pJsn
JJp
sur8rro
leqt
MorIS
VN(
Surler[dar
Surzrrapereqt
ro;
sanbruqrel
snorJpl
aqt taqlaSol
uJ>leJ
'puJ
eqt
JJo
sunJ >lroJ
Juo
sP Jrnlf,nJls-I
e
ol
peua^uoJ
sr elqqnq
eql
WIqM
le lulod
rql
lp
>leerq
e
qlrM
'qled
snonurl
-uoJsrp
e sMolloJ
Jlqqnq
JrJtJrr[u.dse
uy
.selqqnq
Jo sJJnlf,nrts-I
elqnop
yo
sqted
Jqt
JurrurJtJp
suorlprJprsuor
snoSoleuv
'VNe
JeJurl
{uorJ
d1a.,lrsua1xa
lsour
selerlJp
JloJJJJql
eJnlJnrls
Jql
pue
'er8ual
Jrups
eqt
ere
sJqf,uerq
Jrrqt
IIe
ueqM
srnJJo
turod
uortralJul
uV
'qted
snonurl
-uoJ
p
sMolloJ
'tuaru8erl
reJurl
e
Buole
sa.tour
>lJoJ
euo qllqm
ur
'eJnt)nJts-tr
aldurs
y
'JZrs
ur
pJlqnop
]eqt
\/N(I
Jo
Jln)3loru
JeJurl
e,{q
pauolloJ
rq plnoM
teqt
eurl
eql
ruorl uou
-er^ep
rraqt
[q
pJrnseJur
'sqted
JrtsrrJ]Jereq)
,lrolloJ
sJInJJlotu
Surlerrldar
;o
sad,{1
tualJJJr(I
'adeqs
Lq
a.rou peururJlap
sr
Jua{uJloru
JrJqM
uorsueurp puoJJs
e
pue
sselu r(q
saleredas
leql
uorsuJurrp
lsJrJ
e
ur
pasaroqdorlJJle
ere
VNe
uorrloeu
aql
EI
ulLdvHl
Surle:rldar
;o
sluaur8erJ
uorlJrrlsaJ
qJrqM
ur
'anbruq;a1
Surddeu
lpuorsuJrurp-oml
aqt sJlen
-snlll
+':j i
:iiJill..rj
j
'vNo
Jo
uorlerSrur
rrlaroqdorl
-ra1a
uodn
aneq adeqs ur sa8ueqt
lpqt
sDJJJe
aqt
Jo
aSelue^pe
sJ>le] uorlnlosJr
ralearS qlrm
sur8rro
Surdderu JoJ
poqlJru
lueJaJ
JJotrr
V
'seruosoruoJqr
rr1o,{re1na
;o
suorrldar
ur
pJAJJsqo
Llensn
utared
Jql sr srqJ
'a.{a
aqt
Io
spua
ryoq
w
uraned
(letrrlaurruds)
e
sarnpord
uoueJrTdJJ
IeuorDeJrprg
'aLa
aql
;o
pue
au)
le
rsqlo aqt dq
pe1l{olloJ
rq
ol
Ieqpl
Jo
'u0rlrpuol
palelAqlatu
f11n1
aq1 selolsai
aselfq1au up0
aql
lrlun
selrs
llvg
le
alpls slr
surplulpur
qrlqM
'VN6
pa1e1fiqlauLtuaq
sanrb
yg6 pelelAqlaur
1o
uorlerLldeX
,i'i:j i
tH{ti}j:i
'vN0
le0
-urt
p
roJ
qled
eq1 sMoqs
aurt
poqspp
oqI'(aurt
prlos) qled
rLlaroqdotpela
s1L
fq
pamollo1
eq
upl
qlrqM
'1uaLu6erl
uorllulsa.l
6uqerrldai
e
1o
edeqs eql
ourulalap
slro1
6uL1e:r1do.l
Jo
.laqulnu
aql
pup
urbuo
aq1
1o
uorlrsod
aq1
*'-.i:.
jiS*ilJ.jj
08€
<-
sseu Iq
setBtedes uotsueu-ltp
]sltl
qlt
qtr
qlI
,,,*";
ll;
olqqng
A
olqnoc
l
aldurrg