Хэллем Э. Интерпретация фаций и стратиграфическая последовательность
Подождите немного. Документ загружается.


10,
ФАЦИИ
И
БИОСТРАТИГРАФИЯ ФАНЕРОЗОЯ
281
концентрируется
в
абиссальных красных глинах,
В
настоящее
время
мы все еще
практически ничего
не
знаем
о
концентра-
циях иридия
в
других частях стратиграфического разреза,
и
надо провести гораздо больше анализов, прежде
чем
уверенно
утверждать,
что
отмеченные содержания действительно пред-
ставляют собой заметную аномалию.
Еще одна серьезная проблема
для
современных катастрофа
стов возникла после недавнего выступления итальянского гео-
лога Вецеля
[F. С-
Wezcl],
который опровергает мнение
о том,
что
в
классическом разрезе Губбио переход
от
меловых
к
тре-
тичным отложениям непрерывен. Согласно детальным исследо-
ваниям Вецеля, многие осадки, считавшиеся пелагическими,
в действительности являются турбидита«п
с
переотложеннымн
раковинами фораминифер
[456].
Похоже, однако»
что
массовое вымирание
в
конце мела дает
еще один пример того,
что
называют «эффектом Фогта—Хол-
дена»
[33]:
«Новые данные, сколь
бы ни был
надежен
их
источ-
ник
и
СКФЛЬ
бы ни
было высоко
их
качество, почти никогда
не
перечеркивали старых теоретических представлений,
но они
все-
гда служат пищей
для
новых
и еще
более диковинных идей».
Те,
кто не
склонен
к
сарказму
или
пессимизму, могут найти
утешение
в
несколько менее категоричном афоризме Оскара
Уайлда: «Истина никогда
не
бывает абсолютной
и
редко бывает
простой».
ФАУНИСТИЧЕСКИЕ ПРОВИНЦИИ
Используя богатый материал недавно опубликованных работ
о палеобиогеографическом распределении организмов
и
наши
сильно возросшие знания
о
физико-географических условиях
прошлого,
мы
вполне могли
бы
сделать теперь определенные
выводы
о том, что
влияет
на
обособление фаунистичеекпх про-
винций.
Два
наиболее общепризнанных фактора
— это
климат
и движение плит.
Климат
и
колебания
уровня моря.
Поскольку климат несом-
ненно играл определяющую роль
при
формировании фауипсти-
ческих провинций четвертичного периода,
не
удивительно,
что
многие палеонтологи считают
его
главным фактором, влиявшим
на распределение организмов
и в
более отдаленные времена.
Однако
в
мезозое ровный, спокойный климат должен
был
ока-
зывать меньшее влияние
на
развитие фауны,
и
соответственно
его влияние труднее выделить
из
переплетения других факторов
Хорошим объектом исследования
в
этом случае служит юрский
период,
С
одной стороны,
и юре
имеется много данных,
а с
дру-
гой
— в это
время движения плит, несущих
на
себе континенты,
были незначительными; единственным важным событием такого

282
10.
ФАЦИИ И БИОСТРЛТИГРЛФИЯ ФАНЕРОЗОЯ
рода было образование в конце периода узкого океана в цен-
тральной части Атлантики.
Наиболее очевидно важнейшая роль климата проявляется
в распределении организмов по широтам. При этом такие фак-
торы,
как сезонность и дневная освещенность, имеют здесь не
меньшее значение, чем простое изменение температуры. Как
показано в гл. 7, тропические пояса и умеренные климатические
зоны можно выделить и но наземным растениям, и по морским
беспозвоночным. Наиболее интересно выяснить происхождение
и историю развития
тетического
и
бореального
морских поясов
{realms или superprovinces). Их отличительные черты эпизоди-
чески проявлялись еще в ранней юре, но окончательно они
оформились в середине юры и продолжали существовать в ран-
пемеловую эпоху. Детальное описание соответствующих фауми-
стических комплексов приведено в работе автора [187] и
в книге под редакцией Кейзи и Росона [54].
За редкими исключениями, обособление типичной бореаль-
ной фауны, ограниченной северными частями Евразии и Север-
ной Америки, затронуло только аммонитов и белемнитов. В ос-
тальном для бореального пояса в целом характерны меньшая
плотность и пониженное таксономическое разнообразие таких
типичных для пояса Тетис групп, как литуолиды из форамини-
фер,
герматипные кораллы, гидроидные, а из двустворок руди-
сты.
С другой стороны, большинство групп двустворчатых мол-
люсков в бореальном поясе по меньшей мере столь же разно-
образны. Переход от одного пояса к другому был постепенным
и географически несколько менялся во времени.
Наиболее популярна интерпретация, которая состоит в том,
чго бореальная фауна развивалась в водах, слишком холодных
для вторжения фауны пояса Тетис, но такое суждение заклю-
чает в себе и некое противоречие. Как показывает изучение на-
земной флоры того времени, температурные градиенты от тро-
пиков к полярным районам были тогда гораздо меньшими, чем
ныне. И все же юрская аммонитовая фауна заметно менялась
с широтой, так что было совсем немного (а в экстремальных
случаях не было вовсе) родов, общих для обоих поясов, несмо-
тря на то, что морские условия всегда были более однородны,
чем наземные.
Европа представляется наиболее подходящим регионом для
изучения этой проблемы. Поэтому несколько лет назад я попы-
тался исследовать возможную связь распределения юрской
фауны с фациальпымп особенностями этого региона
[184].
Ока-
залось, что такая связь действительно существует для всего
юрского периода, причем для пояса Тетис характерно распрост-
ранение аммонитов главным образом в карбонатных отложе-
ниях, а для бореального пояса — в терригенных фациях
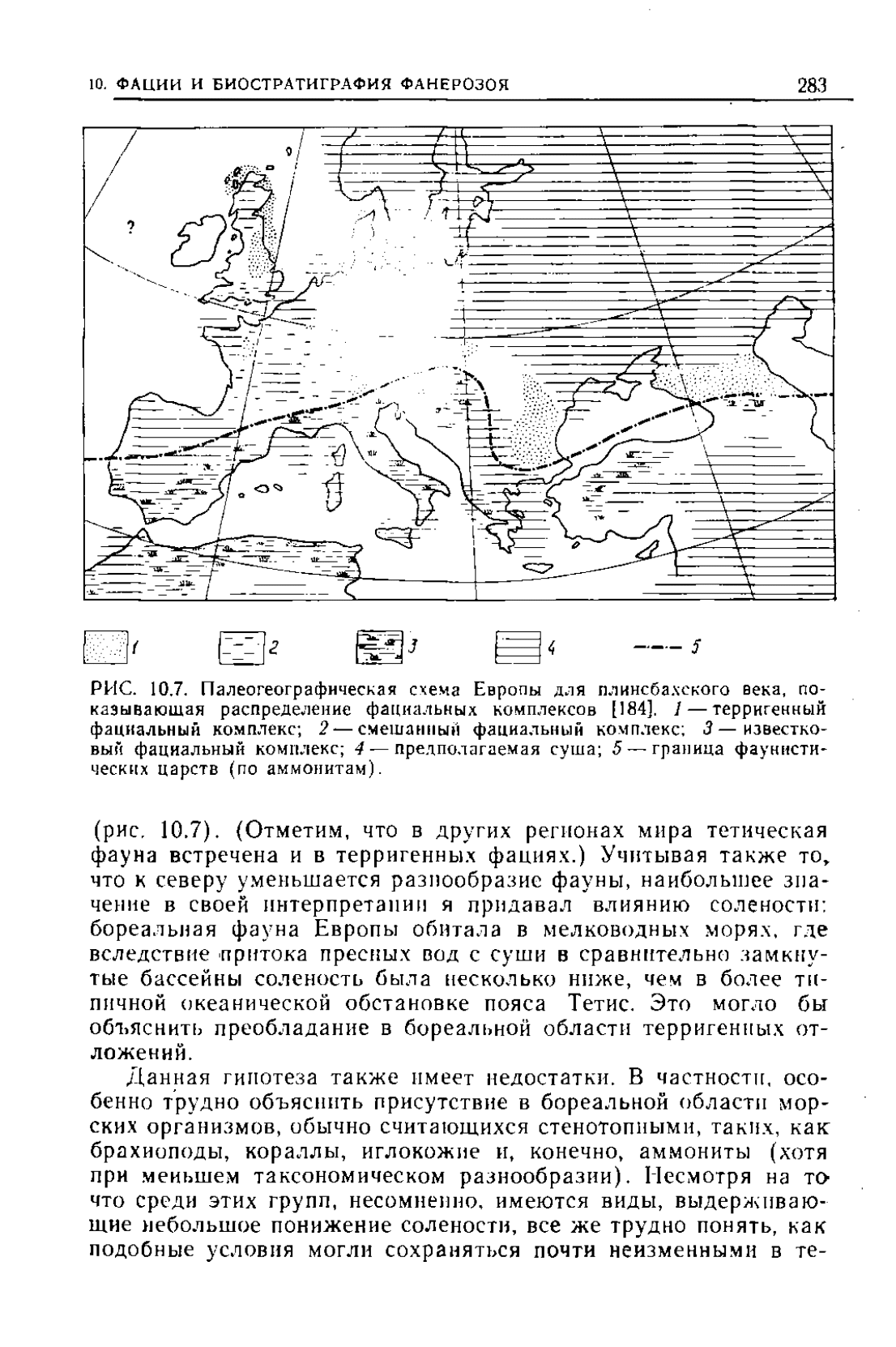
10-
ФАЦИИ И БИОСТРАТИГРАФИЯ ФАНЕРОЗОЯ 283
РИС.
10.7. Палеогеографическая схема Европы для плинсба.\ского века, по-
казывающая распределение фациальных комплексов
[184].
/ — террнгенный
фациальный комплекс; 2 — смешанным фациальный комплекс; 3 — известко-
вый фациальный комплекс; 4 — предполагаемая суша; 5 —граница фауннсти-
ческнх царств (по аммонитам).
(рис, 10.7). (Отметим, что в других регионах мира тетнческая
фауна встречена и в терригенных фациях.) Учитывая также то*
что к северу уменьшается разнообразие фауны, наибольшее зна-
чение в своей интерпретации я придавал влиянию солености;
бореальная фауна Европы обитала в мелководных морях, где
вследствие притока пресных вод с суши в сравнительно замкну-
тые бассейны соленость была несколько ниже, чем в более ти-
пичной океанической обстановке пояса Тетис. Это могло бы
объяснить преобладание в бореальной области терригенных от-
ложений.
Данная гипотеза также имеет недостатки. В частности, осо-
бенно трудно объяснить присутствие в бореальной области мор-
ских организмов, обычно считающихся стенотоппыми, таких, как:
брахиоподы, кораллы, иглокожие и, конечно, аммониты (хотя
при меньшем таксономическом разнообразии). Несмотря на то-
что среди этих групп, несомненно, имеются виды, выдерживаю-
щие небольшое понижение солености, все же трудно понять, как
подобные условия могли сохраняться почти неизменными в те-

284
10.
ФАЦИИ И БИОСТРАТИГРАФИЯ ФАНЕРОЗОЯ
чение довольно долгого времени. К тому же более определенно
выраженные условия солоноватых «рэтеких» лагун очень быстро
сменились в ранней юре — после эветатического поднятия
уровня моря — более нормальными морскими условиями (см.
гл.
5). Поэтому я оставил гипотезу о влиянии одной только со-
лености п стал отдавать предпочтение гипотезе, включающей
концепцию Сандерса [396] об устойчивой связи организмов
с определенными обстановками: тетическая фауна была более
стенотопной и не смогла приспособиться к менее стабильной
или менее закономерно меняющейся обстановке мелководного
эпиконтинентального бореального моря с его колебаниями со-
лености, температуры и, возможно, содержания кислорода [ 187].
Хотя в таком понимании и имеется, вероятно, какая-то доля
истины, все же оно не может служить полным объяснением;
ото стало особенно очевидно после того, как гипотеза Сандерса
о приуроченности организмов к стабильным условиям под-
верглась в последнее время критике [1], При данном подходе
не учитывается, вероятно, палеогеографическая составляющая.
Едва ли можно считать простым совпадением, что разделение
аммонитов на тетические и бореальные было наиболее резким
в батском и тптонско-волжском веках (так что точная корреля-
ция оказалась до сих пор невозможной), которым соответствуют
фазы морской регрессии. Значительное обмеление и уход моря
из обширной части Северо-Атлантического региона произошли
в начале средней юры, а а конце юры регрессия охватила, по-
видимому, весь мир. Наоборот, с трансгрессиями совпадают, как
отмечалось раньше, фазы распространения н радиации фауны.
Рассматриваемую проблему можно изучать дальше на при-
мере двустворчатых моллюсков — одной из наиболее многочис-
ленных и разнообразных групп юрских беспозвоночных. При
анализе их родового состава в мировом масштабе выясняется
четкая обратная корреляция между степенью эндемичпости дву-
створок и площадью современных континентальных областей,
которые в юре были залиты морем, что, вероятно, связано с ко-
лебаниями среднего уровня (рис. 10.8). Существования такой
зависимости следовало ожидать. При низком уровне моря и со-
кращении морских бассейнов миграция фауны вдоль областей
континентального шельфа затруднялась. В результате обмен ге-
нами у малоподвижных организмов, населявших мелководные
районы, становился слабым, что приводило к более локальному
видообразованию.
В гл. 5 достаточно подробно обсуждались доказательства
TOi(J,
что физико-географические условия эпиконтинентальных
морей в ряде аспектов резко отличались от системы открытых
океанов. Можно предполагать, что эти различия и обусловили
развитие эндемичной фауны, .причем степень эндемичности дол-
ч

10. ФАЦИИ H БИОСТРАТИГРАФИЯ ФАНЕРОЗОЯ
285
HSPT A-B Ж 'U 0 К T
РИС. 10.8. Изменение в юрском периоде степени эндемизма родов двуствор-
чатых моллюсков для разных регионов (нижняя диаграмма) и изме-
нение площади современных континентов, покрытых в юре морем,
[189],
На осях абсцисс обозначены века юрского периода; И—геттанг,
S—синемюр, P— плинсбах, Т — тоар,
А—В—
аален и банос, Bt— бат, С —
келловен, О — Оксфорд, К — кнмеридж, T—титон.
жна быть как-то связана со степенью обособления бассейнов.
В верхнемеловых отложениях Западного Внутреннего бассейна
США присутствует эндемичная фауна аммонитов, ассоциирую-
щаяся с фауной других беспозвоночных более ограниченного
разнообразия по сравнению с фауной побережья Мексиканского
залива
[264].
Напрашивается аналогия с пространственно более
распространенной эндемичной бореальной фауной юры — ниж-
него мела. Насколько мне известно, никто не связывал ее появ-
ление просто с влиянием климата. Ниже приведены примеры
эндемизма у обитавших в эпиконтииентальных морях трилоби-
тов.
На важную роль палеогеографического фактора в возник-
новении эндемичной тетической и бореальной фауны указал Ap-
келл [14]. К его мнению присоединились Фюрзнх и Сайке —
авторы работы [149] о расселении моллюсков в Оксфорде на
северо-западе Европы. Вполне могло накладываться действие
и других факторов, например климата, но я подозреваю, что
это имело подчиненное значение. Возможно, более существенное

286
10.
ФАЦИИ И БИОСТРЛТИГРЛФИЯ ФАНЕРОЗОЯ
влияние оказывало взаимодействие различных составляющих
биоты. Когда бореальное фаунистическое сообщество образова-
лось,
оно могло сопротивляться вторжению тетической фауны.
Так, массовая колонизация Северной Европы тетическими ам-
монитами произошла в тоаре — только после того, как вымерло
плинсбахское эндемичное бореальное семейство Amalthldac.
Предложенную зависимость между относительным уровнем
моря и степенью эндемизма необходимо проверить для других
групп ископаемых организмов и геологических систем. По-ви-
димому, альтернативную по отношению к климатической интер-
претацию возрастания степени эндемизма брахиопод от силура
к раннему девону дают Кокс и Мак-Керроу [69], поскольку
эти исследователи связывают данный процесс в основном с рас-
ширением эпиконтинентальных морей (см. гл. 6). И действи-
тельно, как доказывают Джонсон и Буко
[252],
уменьшение эн-
демизма брахиопод от раннего девона к позднему было ре-
зультатом облегчения связей между отдельными регионами по
мере того, как расширялись морские проливы.
К другим примерам по палеозою, очевидно, относятся кем-
брийские трилобиты и пермские фузулипиды. В кембрии при
постепенном расширении эпиконтинентальных морей эндемизм
мелководных трилобитов должен был уменьшаться, а в поздней
перми падение уровня моря должно было привести к повыше-
нию эндемизма у фузулинид, для которых наиболее благопри-
ятной была мелководная среда обитания. В обоих случаях, по
сообщениях Палмера (A. R. Palmer) и Одзавы {Т. Ozawa), из-
менение эндемизма во времени действительно происходило.
Движения плит. Пытаясь установить зависимость между из-
менением положения материков и распределением различных
фаунпстических комплексов, важно отделить влияние фациаль-
ных особенностей. Выделение собственно фаунистичеекпх обла-
стей не должно, насколько это возможно, зависеть от фациаль-
ного состава отложений. Это хорошо иллюстрируется на при-
мере раннеордовикских трилобитов. Применяя статистический
анализ, Уиттингтон и Хьюз [503] выделили следующие четыре
области, которые были затем использованы для реконструкции
континентов:
Область Sclenopeliis (Великобритания, Франция, Марокко, Че-
хословакия, а также —при анализе на уровне семейств —
Китай).
Область Asaphida (Польша, Швеция, Эстония).
Область Bathyurida (Ньюфаундленд, Северо-Восток СССР,
США, Шпицберген, Казахстан).
Область Asaphopsis (Южная Америка, Австралия).
Форти [140] показал, однако, что представители всех обла-
стей,
кроме Selenopeltis, имеются на Шпицбергене, который был

[О,
ФАЦИИ И БИОСТРЛТПГРАФИЯ ФАНЕРОЗОЯ 287
Высопозндемичные
Низкоэндемичные
(провинциальные)
сообщества
/ландемичные)
сообщества
РИС.
10.9. Палеогеографическая модель размещения различных типов сооб-
ществ в раннеордовикском эпикоитинентальном море ([14O]
1
с изменениями).
отнесен VnTTHHrTOHONf и Хьюзом исключительно к области Bat-
hyurida. Фортн смог доказать, что фаунистичеекпе комплексы
ордовика Шпицбергена объединяются в три сообщества, свя-
занные с фациями, каждая из которых интерпретируется как
результат осадконакопления при различной глубине моря
(рис. 10.9). Только в условиях самой мелкой воды развивалось
высокоэндемичное сообщество илленид и хейрурид, поэтому
данное сообщество — одно из наиболее подходящих для иссле-
дования провинциальных особенностей фауны. Аналогичным
образом связано с фациями распределения кембрийских трило-
битов. Вывод об этом сделан Палмером [353] по почти панде-
мичпым агностидам, считавшимися наименее пригодными для
палеобиогеографического анализа.
Такие соотношения можно найти и в других группах и си-
стемах. На ум сразу же приходят хорошо изученные — пред-
положительно глубоководные — сообщества силурийских бра-
хиопод [70]. Точно так же можно показать, что некоторые
группы юрских двустворок, особенно отдельные птериоиды,
имели сравнительно широкое распространение и населяли, как
правило, глубоководные районы морей, тогда как другие, на-
пример гиппуритпды и тригонппды, имел.и относительно много
эндемичных родов
[189].
Учитывая все то, что известно о возможностях расселения
личинок существующих ныне беспозвоночных, я предложил раз-
личать два типа изменения границ фаунистпческнх провинций
со временем применительно к движениям плит
[185].
При
кон-
вергенции
степень сходства фауны различных регионов возра-
стает от более раннего времени к более позднему (рис. 10.10);
при
дивергенции
отмечается противоположное явление.
В табл. 10.2 перечисляется ряд таких примеров, в том числе от-
носящихся и к неморским позвоночным.
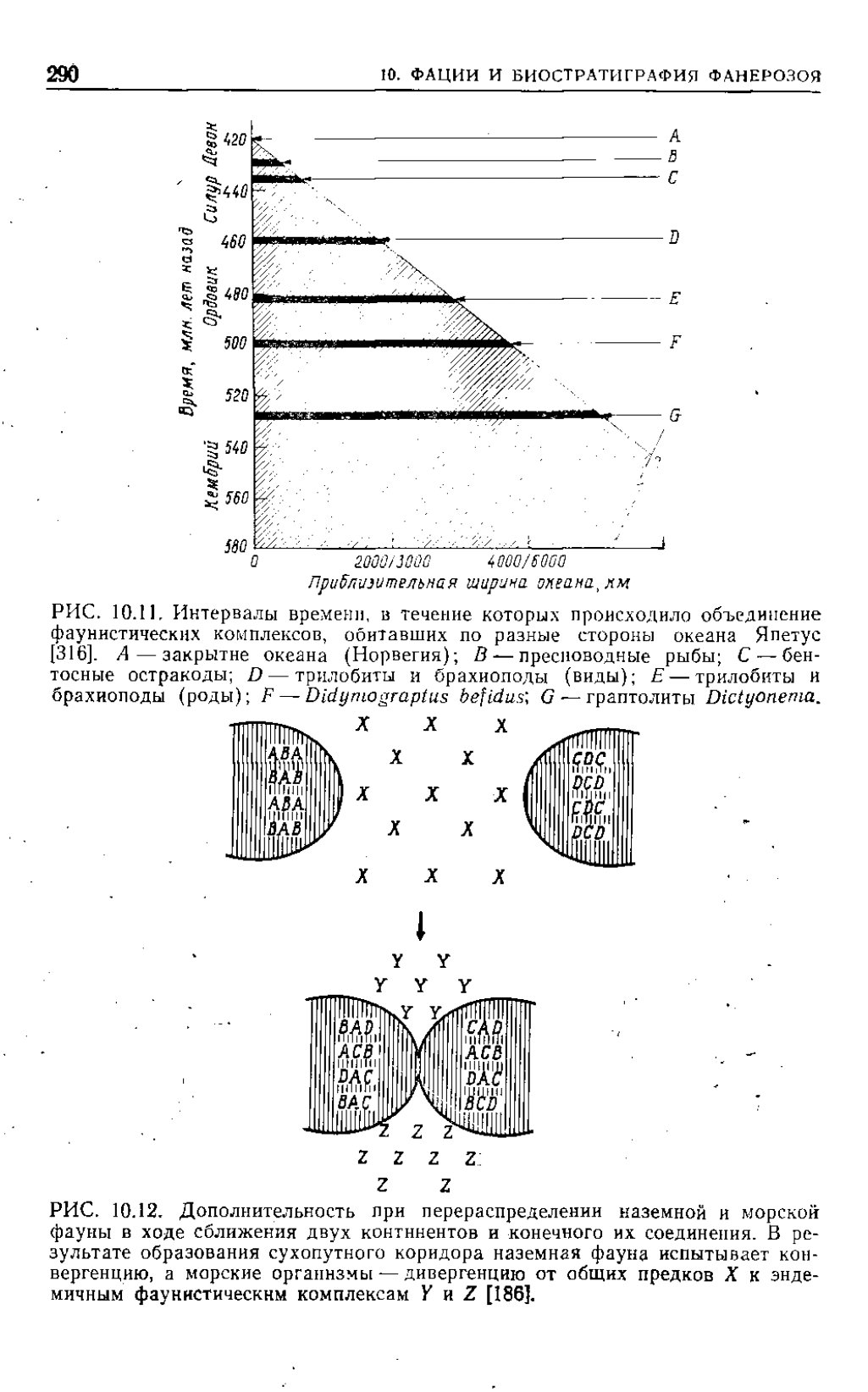
Закрытие в раннем палеозое океана Япетус (существовав-
шего на месте Северной Атлантики) лает особенно хорош и й

288
10.
ФАЦИИ И БНОСТрАТНГРЛФИЯ ФАНЕРОЗОЯ
BAB
ABA
BAB
ABA
BAB
А ВАЗА
DCDC
BABA CDCD
ABAB CDCD
BACDACD
CBADC^
DACDB
ACBAD
CBADCBD
РИС.
10.10, Схема развития конвергенции фаунистических комплексов различ-
ных континентов (заштрихованные квадраты). Когда континенты в резуль-
тате разрастания морского дна приближаются друг к другу, фаунистические
комплексы (обозначенные буквами A
t
б, С, D) постепенно сливаются. У ком-
плексов В и D потенциал миграции выше, чем у комплексов А и С
[186].
пример конвергенции, затронувшей много групп организмов.
Этот пример не связан с влиянием фаций. Полное описание
данных приведено в работе Цпглера и др.
[513].
Мак-Керроу
и Кокс [316] попытались определить относительные скорости
миграции различных фаунистических групп через океан Япетус
if, опираясь на данные о миграции пелагических личинок и
о скорости субдукции, получили оценки изменения ширины
океана со временем. Согласно их интерпретации, пелагические
животные (граптолиты) пересекли океан первыми; за ними по-
следовали животные, у которых в пелагических условиях раз-
виваются личинки (трилобиты, брахиоподы); животные же,
которые в своем развитии не проходят пелагической стадии
(бентосные остракоды), не могли пересечь океан, пока его бе-
рега не сомкнулись хотя бы и не по всей длине, а только в од-
ной точке. Наконец, фаунистические комплексы, обитавшие
в пресной или в солоноватой воде, не могли перейти с одного
континента на другой, пока не появились неморскне перешейки
(рис.
10.11).

IO ФАЦИИ И БИОСТРАТИГРАФИЯ ФАНЕРОЗОЯ
289
ТАБЛИЦА 10.2
СОПОСТАВЛЕНИЕ СОБЫТИИ, ОПИСЫВАЕМЫХ ТЕКТОНИКОЙ ПЛИТ,
И ИЗМЕНЕНИЙ В РАСПРЕДЕЛЕНИИ ФАУНИСТИЧЕСКИХ
КОМПЛЕКСОВ. (Литературные источники приведены в работе автора,
опубликованной в 1974 г. [186])
События, описываемые Фаункстическне комплексы, Фауннстические комплексы,
тектоникой плит испытывающие конвергенцию испытывающие дивергенцию
Закрытие прото-Атлаи-
тического океана (ор
довик, силур)
Закрытие Уральского
морского прохода
Раскрытие Атлантиче
ского океана (мел,
третичный период)
Раскрытие Индийского
океана (мел)
Закрытие океана Тетис
(поздний мел; середи
на третичного перио-
да)
Трилобиты, граптолнты,
кораллы, брахиоподы,
конодонты, апаспнды
и телодонты двух кон-
тинентов по обе сто-
роны прото-Атланти-
ческого океана
Послепермскне конти-
нентальные позвоноч-
ные Евразии
(?) Аммониты Евразии
и Африки — Аравии.
Млекопитающие Евра
зии и Африки
Меловые двустворки л
бентосныс форамини-
феры Карибского и
Средиземного морен,
Позднемеловые аммо-
ниты США и Запад-
ной Европы—Северной
Африки- Послераиие-
эоцеповые млекопи-
тающие Северной Аме-
рики и Европы. Тре-
тичные млекопитаю-
щие Африки и Южной
Америки
Двустворки Восточно-
Африканского и Ин-
дийского шельфов
Моллюски, форамини-
феры и т. д. Индий-
ского и Средиземного
моря — Атлантического
океана
Третью схему биостратиграфического развития я назвал до-
полнительностью
(complementarity). При изменении границ
смежных морских и неморских областей обитания в одной
группе может проявляться конвергенция, а в другой диверген-
ция—это и есть дополнительность. Например, если создается
сухопутное соединение между двумя ранее изолированными кон-
тинентальными областями, становится возможной конвергенция
наземной фауны; и наоборот» в случае раскола некогда единой
19 Заказ № 389
290
10.
ФАЦИИ И БИОСТРАТИГРАФИЯ ФАНЕРОЗОЯ
0
2000/J000 4000/6000
Приблизительная ширина опеана^м
РИС. 10.11,
Интервалы времени, в течение которых происходило объединение
фаунистических комплексов, обитавших по разные стороны океана Япетус
[316].
А — закрытие океана (Норвегия); В — пресноводные рыбы; С — бен-
тосные остракоды; D — трилобиты и брахиоподы (виды); E — трилобиты и
брахиоподы (роды); F —
Didymograptus befidus\
G — граптолиты
Dictyonema
K
Z Z 2 Z
г
z
РИС.
10.12, Дополнительность при перераспределении наземной и морской
фауны в ходе сближения двух континентов и конечного их соединения. В ре-
зультате образования сухопутного коридора наземная фауна испытывает кон-
вергенцию, а морские организмы — дивергенцию от общих предков X к энде-
мичным фаункстическнм комплексам YnZ
[186].