Balian E.V., L?v?que C., Segers H., Martens K. (Eds.) Freshwater Animal Diversity Assessment
Подождите немного. Документ загружается.


collected from lentic environments, b ut at least one-
third of the species are associated with lotic habitats.
For example, the 16 species of Agriotypus are only
found in fast-running streams in Europe and Asia
(Bennett, 2001) and species of the diapriid genus
Psychopria Masner and Garcı
´
a occur in running
water in the new world (Masner & Garcı
´
a, 2002).
Phylogeny and historical processes
The earliest fossil Hymenoptera are herbivorous
sawflies from the late Triassic (230 mya) (Riek,
1955) and they were almost certainly terrestrial, as
are all extant sawfly species. Aquatic development is,
therefore, apomorphic within Hymenoptera and has
evolved at least 50 times based on the number of
unrelated genera of aquatic Hymenoptera (Table 2).
Indeed, the only suprageneric aquatic radiation within
the order may be within several putatively related
genera of aquatic Scelionidae (Johnson & Masner,
2004).
How did aquatic parasitism evolve? It is likely that
it did not evolve de novo from terrestrial parasitism,
but rather through transition from wasps parasitizing
semi-aquatic hosts around the water’s surface .
Females of these semi-aquatic species would benefit
from characteristics that help movement on and
Table 1 Number of species of aquatic Hymenoptera by biogeographic region
PA NA NT AT OL AU PAC ANT World
Superfamily: Chalcidoidea 24 22 7 87 2 – 51
Family: Chalcididae 3 3 - - - - - - 6
Family: Eulophidae 8 3 1 - - - - - 12
Family: Mymaridae 4 4 2 - 1 1 - - 6
Family: Pteromalidae 4 4 - - - - 1 - 5
Family: Trichogrammatidae 5 8 4 - 7 6 1 - 22
Superfamily: Cynipoidea 2 3 – – – – 7 – 10
Family: Figitidae 2 3 – – – – 7 – 10
Superfamily: Ichneumonoidea 29 20 2 1 16 1 – – 65
Family: Braconidae 16 9 – 1 2 – – – 26
Subfamily: Alysiinae 12 4 – – – – – – 16
Subfamily: Braconinae – 1 – – – – – – 1
Subfamily: Microgastrinae 1 – – – 1 – – – 2
Subfamily: Opiinae 3 4 – 1 1 – – – 7
Family: Ichneumonidae 13 11 2 – 14 1 – – 39
Subfamily: Agriotypinae 6 – – – 10 – – – 16
Subfamily: Campopleginae 1 – – – 3 – – – 3
Subfamily: Cremastinae – 6 1 – – – – – 7
Subfamily: Cryptinae 6 4 1 – 1 1 – – 12
Subfamily: Metopiinae – 1 – – – – – – 1
Superfamily: Platygastroidea 2 5 7 – 4 – – – 17
Family: Scelionidae 2 5 7 – 4 – – – 17
Superfamily: Proctotrupoidea 2 3 1 – – – – – 6
Family: Diapriidae 2 3 1 – – – – – 6
Superfamily: Vespoidea 1 – – – – – – – 1
Family: Pompilidae 1 – – – – – – – 1
Total: Hymenoptera 60 53 17 1 28 8 9 – 150
PA: Palaearctic, NA: Nearctic, NT: Neotropical, AT: Afrotropical, OL: Oriental, AU: Australasian, PAC: Pacific and oceanic islands,
ANT: Antartic
Hydrobiologia (2008) 595:529–534 531
123

around the water’s surface. These features, in turn,
would be pre-adaptations once these species moved
into water. There are at least two characteristics of
aquatic Hymenoptera that appear to be adaptive to
life underwater: (1) short, dense pubescence on the
body and wings that allows the wasp to be hydro-
phobic, as well as to maintain a plastron of air around
the body; (2) elongated, strongly curved claws that
allow gripping of the substrate, so that the female
does not float to the surface or be swept in currents
when searching for hosts. Not all aquatic wasps have
either or both of these features, but many do. Specific
aquatic adapt ations of the egg and larva are not
known, perhaps mainl y because most aquatic Hyme-
noptera are endoparasitoids and, therefore, the habitat
where the host develops has little or no effect on the
parasitoid larva. The ectoparasitoid genus Agriotypus
has evolved a unique pupal respiratory filament that
is necessary because water flow in the host pupal
Trichoptera case is compromised once the host is
killed (Bennett, 2001).
Present distribution and main areas of endemicity
Hymenoptera have been collected in almost every
region of the world from the high Arctic to small,
subantarctic Islands. Aquatic Hymenoptera are known
Table 2 Number of genera of Hymenoptera with at least one aquatic species by biogeographic region
PA NA NT AT OL AU PAC ANT World
Superfamily: Chalcidoidea 11 15 5 – 5 5 2 – 18
Family: Chalcididae 1 1 – – – – – – 1
Family Eulophidae 2 3 1 – – – – – 4
Family: Mymaridae 2 4 1 – 1 1 – – 4
Family: Pteromalidae 3 2 – – – – 1 – 3
Family: Trichogrammatidae 3 5 3 – 4 4 1 – 6
Superfamily: Cynipoidea 2 1 – – – – 3 – 3
Family: Figitidae 2 1 – – – – 3 – 3
Superfamily: Ichneumonoidea 12 12 2 1 6 1 – – 21
Family: Braconidae 6 5 – 1 2 – – – 8
Subfamily: Alysiinae 3 2 – – – – – – 3
Subfamily: Braconinae – 1 – – – – – – 1
Subfamily: Microgastrinae 1 – – – 1 – – – 2
Subfamily: Opiinae 2 2 – 1 1 – – – 2
Family: Ichneumonidae 6 7 2 – 4 1 – – 13
Subfamily: Agriotypinae 1 – – – 1 – – – 1
Subfamily: Campopleginae 1 – – – 2 – – – 2
Subfamily: Cremastinae – 4 1 – – – – – 4
Subfamily: Cryptinae 4 2 1 – 1 1 – – 5
Subfamily: Metopiinae – 1 – – – – – – 1
Superfamily: Platygastroidea 1 4 2 – 2 – – – 5
Family: Scelionidae 1 4 2 – 2 – – – 5
Superfamily: Proctotrupoidea 2 1 1 – – – – – 3
Family: Diapriidae 2 1 1 – – – – – 3
Superfamily: Vespoidea 1 – – – – – – – 1
Family: Pompilidae 1 – – – – – – – 1
Order Hymenoptera 29 33 10 1 13 6 5 – 51
PA: Palaearctic, NA: Nearctic, NT: Neotropical, AT: Afrotropical, OL: Oriental, AU: Australasian, PAC: Pacific and oceanic islands,
ANT: Antartic
532 Hydrobiologia (2008) 595:529–534
123
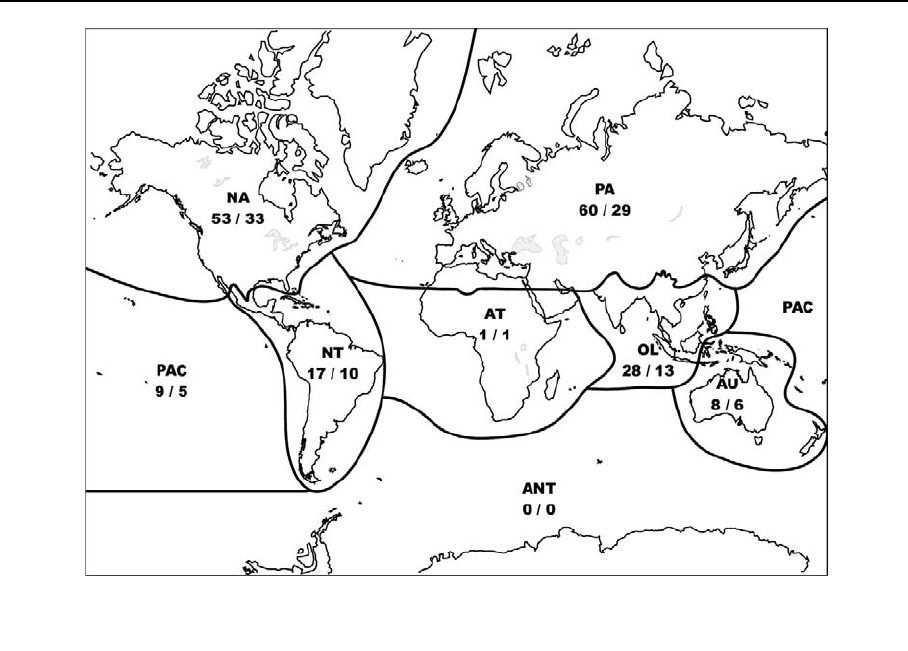
from every major biogeographical region except
Antarctica (Tables 1 and 2; Fig. 2). It is unclear
whether the 150 known aquatic species are a repre-
sentative sample of the total distribution of aquatic
Hymenoptera, because so little is known about the
behaviour, ecology and host range of the vast majority
of species of Hymenoptera. The data in Table 2 are
strongly biased by the amount of study of Hymenop-
tera in each region and extrapolations should not be
made regarding distributional trends of aquatic
Hymenoptera throughout the world. For example,
only one described species of aquatic Hymenoptera is
known from the Afrotropics (the braconid Ademon
angolanus Fischer), but this is likely because of lack
of sampling and biological information for subsaharan
Africa and not because the Afrotropics are actually
devoid of aquatic Hymenoptera. The most noteworthy
area of aquatic wasp endemicity is Hawaii, with
Aspidogyrus and two endemic species of the figitid
genus Kleidotoma Westwood. Aquatic Hymenoptera,
like that of Hymenoptera in general, require much
greater study in terms of collections, rearings and
phylogenetic analyses, which will allow greater
understanding of this fascinating niche and the wasps
that have managed to exploit it.
References
Beardsley, J. W., 1992. Review of the genus Aspidogyrus
Yoshimoto, with descriptions of three new species
(Hymenoptera: Cynipoidea: Eucoilidae). Proceedings,
Hawaiian Entomological Society 31: 139–150.
Bennett, A. M. R., 2001. Phylogeny of Agriotypinae (Hyme-
noptera: Ichneumonidae) with comments on the subfamily
relationships of the basal Ichneumonidae. Systematic
Entomology 26: 329–356.
Bennett, A. M. R., (2007, in press). Aquatic Hymenoptera,
Chapt. 21, In: Merritt, R. W., K. W. Cummins & M. B.
Berg (eds), An Introduction to the Aquatic Insects of
North America. Kendall/Hunt Publ. Co., Dubuque, IA.
Gaston, K. J., 1993. Spatial patterns in the description and
richness of the Hymenoptera. In LaSalle, J. & I. Gauld
(eds), Hymenoptera and Biodiversity. C.A.B. Interna-
tional, Wallingford, UK, 277–293.
Goulet, H. & J. T. Huber, 1993. Hymenoptera of the World: An
Identification Guide to Families. Research Branch of
Agriculture Canada Publication 1894/E, 668 pp.
Fig. 2 Distribution of aquatic Hymenoptera species and
genera by zoogeographical region (species number/genus
number). PA—Palaearctic; NA—Nearc tic; NT—Neotropical;
AT—Afrotropical; OL—Oriental; AU—Australasian; PAC—
Pacific Oceanic Islands, ANT—Antartic
Hydrobiologia (2008) 595:529–534 533
123

Grissell, E. E., 1999. Hymenopteran biodiversity: some alien
notions. American Entomologist 45: 235–244.
von Hedquist, K.-J., 1978. Hymenoptera. In Illies, J. (ed.),
Limnofauna Europaea. Gustav Fischer Verlag, New York,
USA, 286–289.
Jackson, D. J., 1966. Observations on the biology of Caraph-
ractus cinctus Walker (Hymenoptera: Mymaridae), a
parasitoid of the eggs of Dytiscidae (Coleoptera). Trans-
actions of the Royal Entomological Society of London
118: 23–49.
Johnson, N. F. & L. Masner, 2004. The genus Thoron Haliday
(Hymenoptera: Scelionidae), egg-parasitoids of water-
scorpions (Hemiptera: Nepidae), with key to world spe-
cies. American Museum Novitates 3452: 1–16.
Masner, L. & J. L. Garcı
´
a, 2002. The genera of Diapriinae
(Hymenoptera: Diapriidae) in the new world. Bulletin of
the American Museum of Natural History 268: 1–138.
Noyes, J. S., 2002. Interactive catalogue of world Chalcidoidea
2001. CD-ROM. Second edition. Taxapad, Vancouver,
Canada. www.taxapad.com.
Riek, E. F., 1955. Fossil insects from the Triassic beds of Mt.
Crosby, Queensland. Australian Journal of Zoology 3:
654–691.
Roble, S., 1985. Submergent capture of Dolomedes triton (Ara-
neae, Pisauridae) by Anoplius depressipes (Hymenoptera:
Pompilidae). The Journal of Arachnology 13: 391–392.
Stork, N. E., 1996. Measuring global biodiversity and its de-
cline. In Reaka-Kudla, M. L., D. E. Wilson & E. O.
Wilson (eds), Biodiversity II. Joseph Henry Press,
Washington, DC, 41–68.
Ulrich, W., 1999. The number of species of Hymenoptera in
Europe and the assessment of the total number of
Hymenoptera in the world. Polskie Pismo Eno-
tomologiczne 68: 151–164.
Waage, J. & D. Greathead, (eds), 1986. Insect Parasitoids.
Academic Press, London, 398 pp.
Yu, D. S., K. van Achterberg & K. Horstmann, 2005. World
Ichneumonoidea 2004. Taxonomy, Biology, Morphology
and Distribution. CD/DVD. Taxapad, Vancouver, Canada.
www.taxapad.com.
534 Hydrobiologia (2008) 595:529–534
123

FRESHWATER ANIMAL DIVERSITY ASSESSMENT
Global diversity of true and pygmy grasshoppers
(Acridomorpha, Orthoptera) in freshwater
Christiane Ame
´
de
´
gnato Æ Hendrik Devriese
Springer Science+Business Media B.V. 2007
Abstract A small percentage of Orthoptera Acrid-
omorpha is comprised species dependant on
continental water ecosystems. However, as phytoph-
agous insects, they are important at the basis of the
trophic chain, mainly in regions with large permanent
biota resulting from the pulses of the rivers. An
assessment of the composi tion and origin of the
populations from different biogeographical regions is
attempted, and the state of our current knowledge is
given in Tables 1, 2 and 3.
Keyword Acridoidea Tetrigoidea
Adaptation Distrib ution Ecology
Introduction
The Orthoptera are primitive hemimetabolous
insects. They are usually not considered to be aquatic
insects. However, some of their members are some-
how linked to freshwater habitats mainly by a relation
to an aquatic host plant. Species that cannot develop
without freshwater, especially for egg laying and
nymphal development are considered as being
primary inhabitants of freshwater biota. Species
living in or at the border of continental freshwater
habitats, but without being strictly linked to a
dependant plant will only be shortly mentioned.
The fre shwater Orthoptera community is mainly
represented by Acridomorpha Acridoidea. However,
some Tetrigoidea are water dependant, at least at
some stages of their life cycle. Among Ensifera, the
predator Katydid genus Phlugis specialises on
nymphs of aquatic grasshoppers in South America.
Tetrigoidea
Most species of Tetrigoidea can be considered as
limno-terrestrial. However, some groups are clearly
dependent on water: they dive under water, swim very
well and feed on aquatic algae. Since the life cycle of
only a few species (perhaps 1%) of Tetrigoidea is
known—most of them from Europe—it is currently
extremely difficult to assess the degree of water
dependency of this group.
Guest editors: E. V. Balian, C. Le
´
ve
ˆ
que, H. Segers &
K. Martens
Freshwater Animal Diversity Assessment
Electronic supplementary material The online version
of this article (doi:10.1007/s10750-007-9132-z) contains
supplementary material, which is available to authorized users.
C. Ame
´
de
´
gnato (&)
Departement Syste
´
matique et Evolution, Museum
National d’Histoire Naturelle, Entomologie, 45 rue de
Buffon, Paris 75005, France
e-mail: amedeg@mnhn.fr
H. Devriese
Institut Royal des Sciences Naturelles, Entomologie,
29 rue Vautier, Bruxelles 1000, Belgium
123
Hydrobiologia (2008) 595:535–543
DOI 10.1007/s10750-007-9132-z

Acridoidea
South America and Asia have the most important
large river systems with considerable variations in
water levels (10 m) which generate immense surfaces
of freshwater biota. Especially in tropical South
America, rich aquatic radiations of Acrid oidea have
been developed that are not found elsewhere. These
radiations include insects living on:
– floating plants, attached or not to the bottom (two
families and four genera);
– grasses and related plants species adapted to
flooded zones (several tribes);
– palm trees specific of these biota (inundated
tropical forest: Var zea, Igapo) (one subfamily);
– margins of inundation zones and shores of
running waters, but with no other special need
except for a link to the vegetation (marginal
species of different groups). The Oriental and
Afrotropical regio ns are less studied, but the
fauna appears to be quite similar in diversity to
the American fauna. On the other hand, the sub-
Saharan region seems to be poorer; this is also the
case for the palearctic fauna, which comprises
only few species inhabiting marshes, moreover
often not strictly linked to these ecosystems.
Adaptations
The morphology of subaquatic species does not
differ markedly from the general morphology of the
group, with the exception of particular adaptations
in some structures. These adaptations include a
general morphology with a fusiform habitus for
species living only in water and that are used to
swim under the water (Fig. 1A). These species also
developed a strong hind femora, expanded hind
tibiae, including their spines and modified spurs
(Fig. 1B). In some species, the presence of dense
hairs on the margins of the tibiae also help to
reinforce their efficiency to the aquatic habitat. Th is
modification of the hind tibiae is more or less
generalised within all the groups linked to water, but
varies from weakly to strongly developed in the
most aquatic species (Ame
´
de
´
gnato, 1977; Carbonell,
1957; Roberts, 1978). Another adaptation is a
morphological and chromatic mimicry to the plant
habitus, often to Gramineae. Linked to physiology
and etho logy, occurs a modification of the ovipositor
valves. Usually, grasshoppers’ ovipositors are
shaped to burrow in the soil. Water inhabiting
grasshoppers have to oviposit within or at the
surface of plants. Epiphylle egg lying is linked to a
strong regression of the valves, while the endophy tic
egg lying, in the stems of the plants is linked to
cutting edges, and depending on the behaviour of
the species, it is also linked to different kinds of
modifications of the upper and lower surfaces of the
valves, modified to rasp the plant tissues.
Diversity of Orthoptera
The number of Orthoptera species is estimated to be
over 25,000 (OSF2). They are distributed over all
geographic zones, except Antarctica . The suborder
Acridomorpha comprises Acrididea and Tetrigidea.
Fig. 1 (A) Habitus of Marellia remipes (Pauliniidae) ( B)
Specialisation of hind tibia and tarsus in Marellia remipes
536 Hydrobiologia (2008) 595:535–543
123

For the Tetrigidea 1,400 species have been described
in 250 genera (OSF2).
Among Acrididea, only the superfamily Acridoi-
dea is actually known to include families or
subfamilies involved in the aquatic habitats. A recent
estimate o f the Acridoidea mentions 7,000 species in
1,500 genera (Otte, 1995a, b). With regard to this
latest list, the groups concerned are the families
Pauliniidae (neotropical) and Acrididae (world wide) .
It is within this last family that most of the genera
with subaquatic adaptation have evolved, through
several subfamilies, the most important being the
neotropical Leptysminae and the Afrotropical and
Oriental Oxyinae, Tropidopolinae and Hemiacridi-
nae. Most Tetrigoidea groups —currently divided into
nine subfamilies and one unassigned tribe (OSF2),
but in strong need for a suprageneric revision—
comprise limnoterrestrial species in all geographic
zones, but only one, Scelimeninae, has true aquatic
species, which occur in the Oriental region.
Phylogeny and historical process
Tetrigoidea
The oldest taxa that have been attributed to the
Tetrigidae are Archaeotetrix and Prototetrix, from the
lower Cretaceous of Transbaikalia (Sharov, 1971).
These taxa have long elytra, a character that is not
present in extant Tetrigidae. It is, therefore, not sure
that these taxa belong to the same family. Apart from
a poorly described taxon, Succinotettix, from the
Baltic amb er (Piton, 1938), no fossils of the family are
known. Provisional analysis of the phylogeny indi-
cates that the most primitive taxa have a leafl ike
pronotum with a split median carina (Devriese,
unpublished). Such species occur on moist spots, but
seldom near rivers. They have evolved into different
lineages, of which the most aquatic are long-winged
and have expanded hind tibia with reduced spines.
Acridoidea
Orthoptera of acridomorphoid type are known from
the Carboniferous. However, extant groups are recent
and do not antedate the Cenozoic (Ame
´
de
´
gnato,
1993). As they are phytophagous insects, the coevo-
lution with plants has led to a high degree of
specialisation. Thus, in the whole group, ecology and
phylogeny are narrowly linked, and groups associated
to freshwater environments appear to be the result of
such adaptive radiations. It is, especially, striking
in the neotropics. Within Old world Acrididae,
phylogenetic relations are not entirely clear.
The heterogeneity and bad definition of the
subfamily Hemiacridinae (type genus very different
from most of the other genera assigned to ‘‘Hemi-
acridinae’’), is associated to the instability in the
different sources of the lists provided for the Oxyi-
nae, Tropidopolinae and ‘‘Hemiacridinae’’. As a
result, the boundaries among these three subfamilies,
in need of phylogenetic clarification, are imprecise.
Thus, a com plex OTH (Oxyinae, Tropidopol inae,
Hemiacridinae) is used in the illustrations. However,
Oxyinae, as well as some water dependant
Hemiacridinae (inconstantly classified as Oxyinae),
and some Tropidopolinae, could be closely related. In
the Oriental and Afrotropical regions, water depen-
dant grasshoppers could thus also result from a single
radiation. However, phylogenetic studies and field
work needs to be carried, in order to test this
hypothesis.
At the present state of our knowledge, none of the
groups mentioned are closely related to New world
subfamilies. However, the morphological conver-
gence is striking, as well for forest biota than for
grassland biota.
Except for the Copiocerinae of palm trees, which
could be distantly related to the wide-ranging Cyrta-
cantacridoid group, the bulk of Acrididae of humid
biota belongs to only one subfamily, the Leptysminae,
with two main radiations, linked to the main ecosys-
tems: one mostly in forest biota, the Tetrataeniini, and
the other in the grassland biota, the Leptysmini. It is
clear that the two tribes originated in South America.
The subfamily itself is part of a group of subfamilies of
Acrididae also including the Ommatolampinae and the
Rhytidochrotinae (Ame
´
de
´
gnato, 1977), which have
radiated in most environment s.
Paulinia and Marellia, two monospecific genera,
either considered as two subfamilies of Acrididae
(Eades, 2000), partly incertae sedis (Carbonell,
2000), or gathered for practical reason in the same
group Pauliniidae (Ame
´
de
´
gnato, 1974; Ame
´
de
´
gnato,
1977; Dirsh, 1961; Dirsh, 1975; Otte, 1995) are not
closely related (Ame
´
de
´
gnato, 1977; Carbonell, 2000).
They
only
seem to be closer to the Acrididae than to
Hydrobiologia (2008) 595:535–543 537
123
another family of Acridomorpha (Ame
´
de
´
gnato, 1977;
Carbonell, 2000; Eades, 2000), but it is sure that they
are not true actual Acrididae. Their high degree of
divergence is p robably linked to their narrow adap-
tation in a conservative biota, which allowed the
survival of ancient phyla with no more relatives in the
actual fauna, the reason why they are not related to
any other subfamily.
A true continental fauna is characterised by its
composition made of structured groups and subfami-
lies, all linked to each other in coherent entities, as the
result of the biodiversity evolution. So, the insertion of
heterogeneous elements (well-known paraphyletic
Pauliniidae) into a coherent one (neotropical phyla of
Acrididae) is only hampering the understanding of
evolution. These two genera will continue here to be
considered under the same name ‘‘Pauliniidae’’.
Present distribution and main areas of Endemicity
(Fig. 2, 3)
Tetrigoidea (Tables 1, 2, 3)
Among the Tetrigoidea, only the ‘‘Scelimenae verae’’
(sensu Gu
¨
nther, 1938) seem, to be truly aquatic. They
dive under water to hide and to feed on algae that
grow on the underside of boulders (Bhalareo &
Paranjape, 1986; Blackith & Blackith, 1987; Black-
ith, 1988; personal unpublished observations).
However, an examination of the gut content of one
species revealed few aquatic food (Reynolds et al.,
1988). They consist of 79 species that occur in the
Oriental and Australian regions, with a few also
present on the Southeastern border of the Palearctic
region.
Many other species are limno-terrestrial , and
some have adaptations to swimming and are found
at the margins of rivers and lakes. The Neotropical
genera Amorphopus, Eomorphopus and Cr imisus,
the Oriental genera Criotettix, Eucriotettix and the
Afrotropical Morp hopus and Afrocriotettix all occur
on very wet places adjacent to water bodies. In
Europe, Tetrix tu
¨
erki is only found in wet-river
valleys (Nadig, 1991; personal unpublished obser-
vations). But although all these species are
encountered near rivers and lakes, it is not sure,
whether they can be considered to be water
dependent. In Western Africa, most species of the
subfamily Tetriginae occur in seasonaly inundated
grassland (Roy, 1982), but they are also found
elsewhere.
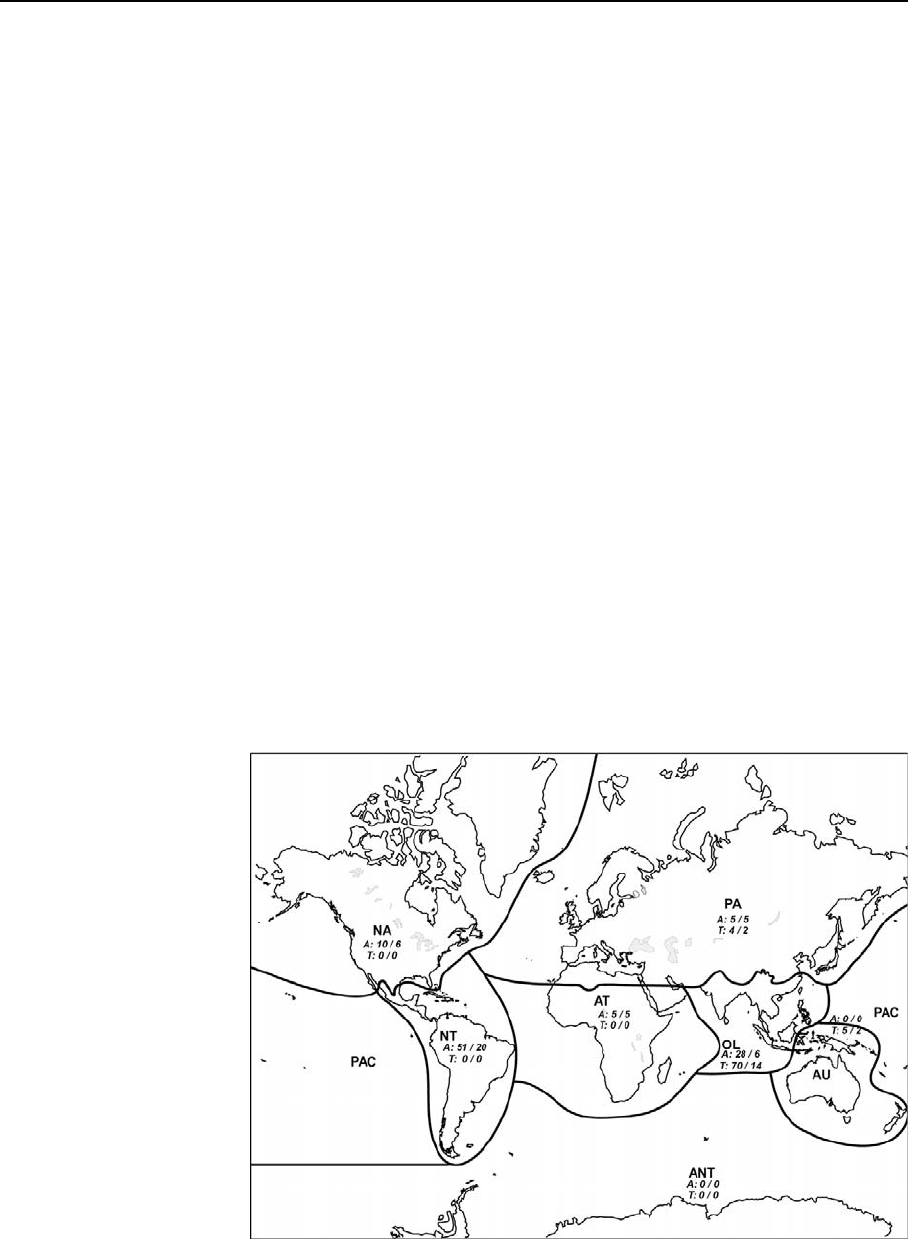
Fig. 2 Distribution of
freshwater (aquatic + water
dependant) Orthoptera
Acridoidea (A) and
Tetrigoidea (T) species and
genera by zoogeographical
region (species number/
genus number). PA,
Palaearctic Region; NA,
Nearctic Region; NT,
Neotropical Region; AT,
Afrotropical Region; OL,
Oriental region; AU,
Australasian Region; PAC,
Pacific Region Oceanic
Islands
538 Hydrobiologia (2008) 595:535–543
123

Acridoidea (Tables 1, 2, 3, Fig. 3)
In the Afrotropical and Oriental regions, where there
are very few studies on aquatic Orthoptera, three
subfamilies of Acrididae are involved in the coloni-
sation of the marshy zones. They belong mainly to
Oxyinae, Hemiacridinae and Tropidopolinae. In this
region, Acridids of the main water surfaces are not
known, with the exception, in India and Southwest
Asia of Gesonula punctifrons (Oxyinae), which is
described as the most aquatic species of Acridoidea,
with egg pods deposited within Water Hyacinth, on
which it also feeds (Kapur & Dutta, 1952; Sank aran,
1976; Sankaran et al., 1966). The inundation zones
seem to be occupied by six other Oxyinae (Oxyini)
and their related groups (members of the subfamilies
Tropidopolinae and Hemiacridinae), but little is
known about the life history of the species , and the
species associations can hardly be recognised from
the literature. For Oxyini, a total of five genera and 23
species are involved (Gesonula punctifrons, Hygra-
cris palustris; Oxya and Oxyina: 18 species; Quilta:
three species (Hollis, 1971, 1975).
The genus Oxya, the main component, extends
over the whole paleotropical region but only one
species occurs in Africa (Oxya hyla). The same is true
for the Hemiacridinae Hieroglyphus: 10 species
mostly Asiatic with only one hygrophilous species
(H. daganensis) in sub saharian Africa. On the
contrary Mesopsera filum (Hemiacridinae) is only
known from central Africa (Uvarov, 1977). Also
mentioned for Africa are some membe rs of the
Tropidopolinae ( Tropidopola nigerica) and another
Hemiacridinae, Spathost ernum pygmaeum in subsah-
arian Africa (Phipps, 1966, 1970). It is clear that the
diversity is centred on India and South western Asia
and that it is sharply decreasing towards Australia
(one species) but also towards Africa. On the other
hand, while the Asiatic fauna is represented by true
hygrophilous Oxyiinae, such forms are poorly repre-
sented in Africa (five species, distributed in five
genera and three subfamilies) (Phipps, 1970). Other
species mentioned in the literature for Africa (COPR
1982; Fishpool & Popov, 1984; Uvarov, 1977) b elong
mostly to groups mainly adapted to other ecosy stems:
Eyprepocnemidinae with Cataloipus, Phyllocercus and
Eyprepocnemis,AcridinaewithDuronia chloronata,
Orthoctha grossa, Jasomenia sansibarica (Phipps,
1966), Calephorus compressicornis, Paracinema
Table 1 Number of Freshwater Orthoptera species per zoogeographic region (Total FW, water dependent plus aquatic or subaquatic species; Wdpdt, Water dependent species).
PA, Palaearctic Region; NA, Nearctic Region; AT, Afrotropical Region, NT, Neotropical Region; OL, Oriental Region; AU, Australasian Region; PAc Pacific Region and
Oceanic Islands
Species number PA NA NT AT OL AU + PAC World
Total FW WDpt Total FW WDpt Total FW WDpt Total FW WDpt Total FW WDpt Total FW WDpt Total FW WDpt
ACRIDIDAE 5 5 10 10 52 48 14 14 28 27 107 102
Leptysminae 3 3 39 35 41 37
Copiocerinae 10 10 10 10
Oxyinae Hemiacridinae Tropidopolinae 5 5 28 27 32 31
Other subfamilies 5 5 7 7 3 3 9 9 24 24
PAULINIIDAE 2 2
Acridoidea 5 5 10 10 54 48 14 14 28 27 109 102
Tetrigoidea 4 4 70 5 5 79 9
ORTHOPTERA 9 9 10 10 54 48 14 14 98 27 5 5 188 111
Hydrobiologia (2008) 595:535–543 539
123

tricolor and Mesopsis abbreviatus. The poverty of the
fauna seems to be real, but it cannot be excluded that, in
Africa, flood plains have not been sufficiently studied,
notably those of the central basin of Congo. Also, the
localisation of the others inundation zones in highlands
or in desertic environments is probably determinant in
hampering the ecological diversification.
The Palearctic and Nearctic zones have closely
related and poor faunas. The Palearctic region, close
to the Oriental one, suffered a drastic impoverish-
ment, notably in Oxyiinae. The groups involved are
of wide distribution, mainly euryapt and are only
marginally adapted to freshwater habitats, by few
species. It is the case for:
– the Melanoplinae (nearctic bush subfamily) with
two genera and several species: Gymnoscirtetes
pusillus, Paroxya clavuliger, P. atlantica, in the
South east of the USA (Squitier & Capinera,
2002) and Paroxya hoosieri, in the great lakes
region (Bright, 2006);
– the Gomphocerinae (a world wide ranging sub-
family of grassland insects) with the genus
Stethophyma, adapted to marshes and peat bogs
Table 2 Number of freshwater orthoptera genera (water
dependant plus aquatic or subaquatic species). PA, Palaearctic
Region; NA, Nearctic Region; AT, Afrotropical Region; NT,
Neotropical Region; OL, Oriental Region; AU, Australasian
Region; PAc Pacific Region and Oceanic Islands
GN: Genera Number PA NA NT AT OL AU + PAc Total
ACRIDIDAE 5 6 18 5 6 34
Leptysminae 3 15 15
Copiocerinae 3 3
Oxyinae Hemiacridinae Tropidopolinae 5 6 9
Other subfamilies 5 3 7
PAULINIIDAE 2 2
Acridoidea 5 6 20 5 6 36
Tetrigoidea 2 14 2 14
ORTHOPTERA 7 6 20 5 20 2 50
Table 3 Taxonomic origin of water dependant Acridoidea and
Tetrigoidea: a biogeographic perspective within the spectrum
of the group diversity. Total = FW + Marine + terrestrial
species, FW = Aquatic/subaquatic + water dependent. Species
numbers are widely provisional; the source is Otte ( 1995a, b);
For Acridoidea, the Ommexechidae, in the Neotropical region
are not included, as well as some small groups in fact
belonging to Pamphagoidea, in all continents
Species number PA NA NT AT OL AU + PAC World
Total FW Total FW Total FW Total FW Total FW Total FW Total Fw
Acrididae 596 5 797 10 1083 52 1459 14 1776 28 572 6283 107
Leptysminae 3 3 80 39 80 41
Copiocerinae 65 10 65 10
Oxyinae, Hemiacridinae Tropidopolinae 42 117 5 58 28 106 323 32
Other subfamilies 554 5 794 7 938 (3) 1342 (9) 1718 ? 466 5812 24
Pauliniidae 2 2 2 2
Acridoidea 596 5 824 10 1539 54 1459 14 1776 28 572 6766 109
Tetrigoidea
a
140 4 20 0 195 0 200 0 700 70 145 5 1400 79
ORTHOPTERA 736 9 844 10 1734 54 1659 14 2476 98 717 5 8166 188
a
N.B. The number of species occurring on the edge of the Palaearctic and the Oriental region in China being substantial, the figures
for both regions have been based on the number of species in resp. predominantly Palearctic (e.g., Tetrix and Formosatettix) and
Oriental regions
540 Hydrobiologia (2008) 595:535–543
123