Александров Ю.И. Основы психофизиологии
Подождите немного. Документ загружается.

на формирование нового. В зависимости от условий, это влияние может быть как
позитивным, так и негативным, т.е. наличие прошлого опыта может способствовать или
препятствовать приобретению нового [Эксперим. психол., 1963]. Возможность
эффективного использования предшествующего опыта при решении новых задач
особенно ярко проявляется в научении принципам решения задач или, как его назвал Г.
Харлоу, – научении обучению [Harlow, 1949]. Резкое увеличение скорости решения в ряду
однотипных задач было обнаружено Г. Харлоу не только у человека, но и у животных.
Таким образом, для современных теоретических представлений о научении характерно
его рассмотрение прежде всего как процесса приобретения нового опыта поведения и
соответствующих ему внутренних ментальных репрезентаций.
2. ПОДХОД К НАУЧЕНИЮ КАК ПРОЦЕССУ
Повышенное внимание к анализу научения как процесса происходит из обыденной задачи
научить, передать опыт. Эта процессуальная сторона научения подчеркивается в его
определениях как в биологии [Хайнд, 1975], так и в психологии [Александров,
Максимова, 1997]. Она же является основой нейробиологического понимания
пластичности нейрональной активности в качестве механизма научения [Котляр, 1989].
В экспериментальной психологии и биологии был разработан ряд модельных ситуаций,
воспроизводящих основные типы ситуаций реального научения. Эти экспериментальные
модели научения перечислены и подробно рассмотрены в руководствах по
экспериментальной психологии [Эксперим. психол., 1963; Эксперим. психол., 1973 и др.]
и обзорах по биологии обучения [Хайнд, 1975; Gould, 1986 и др.]. Среди них можно
выделить ряд наиболее общих типов моделей: классическое обусловливание, оперантное
обусловливание, выработка дифференцировок, обучение в лабиринтах, разрешение
проблемной ситуации, а также специфически человеческие формы вербального научения,
научения путем инструктирования и научения по примеру.
Исследование процессуальной стороны научения в психологии наиболее ярко проявилось
в построении кривых обучения решению разных типов задач. Как правило, это
зависимости эффективности решения задачи (оцениваемой через объем воспроизводимого
по памяти материала либо через количество правильных ответов или, наоборот,
количество ошибок) от количества повторений экспериментальной ситуации. Подобный
анализ позволил выявить фазы, во время которых протекает научение, и описать
динамику научения с помощью математических уравнений [Эксперим. психол., 1963]. В
ряде случаев на кривых обучения наблюдались участки плато, т.е. периоды, когда
улучшения поведения по формальным показателям не происходило. Наличие подобных
участков позволило высказать предположения об изменении стратегий научения в ходе
этого процесса.
Для всех тех форм научения, которые исследовались с помощью построения кривых
обучения, характерно плавное улучшение выполнения задачи. В то же время в некоторых
моделях обучения, в основе которых лежит создание проблемной ситуации (например,
научение в проблемной клетке [Эксперим. психол., 1973]), было обнаружено явление
резкого, скачкообразного изменения хода кривой научения, соответствовшего
нахождению решения задачи с последующим успешным повторением найденного
решения. Это явление получило название инсайта, или озарения.
Инсайт наблюдается в более сложных формах научения по сравнению с условными
рефлексами и дифференцировками. Решение задач, требующих подобных форм научения,
происходит путем проб и ошибок. Предполагается,что инсайт возникает тогда, когда
организм способен прогнозировать результат пробы [Эксперим. психол., 1963], в
противном случае научение протекает плавно.
Исследование научения как процесса, таким образом, позволило выявить два
принципиально разных типа научения: плавное научение и скачкообразное научение
(инсайт и импринтинг). Эта разница может быть связана с разной реорганизацией опыта в
этих ситуациях, однако ответить на вопрос, какой именно, можно лишь выяснив, какие
именно элементы опыта участвуют и формируются в данных формах научения.
3. ПРЕДСТАВЛЕНИЕ О
НЕЙРОФИЗИОЛОГИЧЕСКИХ МЕХАНИЗМАХ
НАУЧЕНИЯ
С появлением методов регистрации активности мозга и особенно активности нервных
клеток проблема научения стала интенсивно исследоваться в нейробиологии. В силу того,
что эти экспериментальные исследования проводились на животных, и из-за трудностей
регистрации нейрональной активности в свободном поведении, основное внимание
нейрофизиологов было сосредоточено на исследовании самых простых форм обучения,
таких, как привыкание, выработка рефлексов и дифференцировок. Находясь под сильным
теоретическим влиянием бихевиоризма, нейрофизиологи ограничивались изучением форм
нейрональной пластичности как основы формирования связей при подобных формах
научения.
В рамках данного направления много сил было потрачено на поиск места локализации
следа памяти (или энграммы), который, как предполагалось, фиксирует образовавшуюся
при научении временную связь. Однако анализ динамики активности отдельных нейронов
в ряду сочетаний стимуляции и подкрепления показал, что изменения импульсации
нейронов происходят практически одновременно во многих отделах мозга, причем
изменения в специфических анализаторных и моторных областях могут наблюдаться
позже, чем в ассоциативных, таких например, как гиппокамп [Рабинович, 1975; Thompson,
1976]. Таким образом, было показано, что в научении задействованы многие структуры
мозга, а не только специфические сенсорные, моторные и связывающие их ассоциативные
структуры, как следовало из классических представлений о замыкании временных связей.
Это вовлечение в процесс обучения многих структур привело к формированию
представления об обучающихся нервных сетях (см. напр. [Eccles, 1977]).
В связи со спецификой нервной ткани, выражающейся в наличии у нервных клеток очень
длинных отростков, способных проводить электрические импульсы, и с представлением о
потоках информации внутри мозга, ключевым механизмом пластичности нейронов, по
мнению авторов, является изменение эффективности синаптической передачи, т.е.
изменения этих информационных потоков. Именно поэтому на субклеточном уровне
большинство исследований механизмов научения заключалось в изучении
закономерностей функционирования синапсов. Этот подход привел к обнаружению
долговременной посттетанической потенциации. Такая форма изменения
функционирования клеточных контактов рассматривается авторами как соответствующая
сформулированным Д.О. Хеббом [Hebb, 1949] принципам синаптической пластичности,
способной обеспечить длительное
сохранение результатов научения.
В силу особенностей уже упоминавшейся бихевиористской позиции нейрофизиологи
исследовали научение как процесс дифференцировки и запоминания значимых стимулов
среды. Для теоретического описания этого процесса были предложены разнообразные
инструктивные теории, описывавшие фиксацию нервной системой навязываемых средой
закономерностей.
Чисто инструктивные представления о научении, для которых характерно признание
полной зависимости научения от закономерностей внешней среды, встречаются довольно
редко. В основном на подобных позициях стоят исследователи, пытающиеся
смоделировать работу нейронных сетей [Spinelly, 1970]. Их модели в соответствии с
разными сочетаниями стимулов («условных» и «безусловных») демонстрируют
определенную динамику состояний, которая и понимается как механизм обучения в
нейронной сети. Однако, как было отмечено еще на заре исследований научения, в
зависимости от мотивации, степени готовности и прошлого опыта научение может
происходить по-разному [Эксперим. психол., 1963].
Большой популярностью среди нейрофизиологов пользуются инструктивно-селекционные
теории. В соответствии с одной из таких теорий, предложенной Экклсом [Popper, Eccles,
1977], обучение происходит в нейрональной сети за счет изменения эффективности
отдельных синаптических контактов, выбор которых производится за счет
инструктирующего возбуждения других синапсов. Эта теория учитывает влияние
мотивации и состояния готовности, описывая их как модулирующее возбуждение
определенных входов нейрона. Однако инструктивным теориям присущ ряд недостатков,
не позволяющих использовать их для описания сложных форм научения (см. гл. 14).
4. СПЕЦИФИКА ПСИХОФИЗИОЛОГИЧЕСКОГО
РАССМОТРЕНИЯ НАУЧЕНИЯ
Психофизиология, в отличие от нейрофизиологии, принимает в расчет субъективное
содержание объективных процессов и, более того, изучает как субъективное соотносится
с процессами жизнедеятельности. В соответствии с решением психофизиологической
проблемы в рамках системной психофизиологии (см. гл. 14) психическое связано с
протеканием системных процессов организации активности мозга в поведении [Швырков,
1978]. С этих позиций отнюдь не все исследования научения в нейрофизиологии можно
назвать психофизиологическими, а лишь те, которые проводятся на бодрствующем
животном и исследуют изменения нейрональной активности в связи с изменением
поведения. В соответствии с этим подходом, собственно психофизиологическими
теориями научения можно считать лишь те теории, которые рассматривают формы и
динамику организации мозговой активности в процессе научения. Как правило, такие
теории оперируют с понятиями нейронных сетей или ансамблей, понимая последние как
набор совместно активирующихся клеток [Dudai et al., 1987]. Как отмечается ведущими
учеными в этой области, «понимание функционирования таких ансамблей может
потребовать дополнительных теоретических и экспериментальных приемов по сравнению
с теми, которые требуются для изучения более простых типов нервной организации»
[Dudai et al., 1987, р. 399].
Таким образом, научение с позиций психофизиологии может быть определено как
формирование пространственно-временной организации активности мозга,
обеспечивающей выполнение приобретаемого в процессе обучения нового поведения и
соответствующей новому состоянию субъекта поведения. При таком определении
становится очевидным отличие психофизиологического подхода к научению от подходов,
характерных для психологии и биологии.
Психофизиологическая теория научения должна описывать взаимодействие организма и
среды в процессе научения и отражение этого взаимодействия и его результатов в
изменениях организации мозговой активности. В соответствии с различными
представлениями о движущих силах научения все психофизиологические теории могут
быть разделены на три типа: инструктивные, инструктивно-селективные и селективные.
В отличие от упомянутых ранее инструктивных теорий, предполагающих изменение
функций нейрона за счет изменения состояния синаптических контактов, селективные
теории научения исходят из существования заложенного в процессе созревания
разнообразия нейронных интеграции и выбора необходимой интеграции в результате проб
во время обучения. Появление этих теорий, получивших распространение сравнительно
недавно [Changeux et al., 1984; Edelman, 1989], связано как с обнаружением врожденных
предрасположенностей к овладению определенным опытом, так и со стремлением
подойти к научению с общебиологических позиций (как к эволюционному процессу).
Подобный подход проявляется в широком использовании знаний об организации работы
иммунной системы в качестве аналогии для рассмотрения процессов формирования
нейронных систем, которые обеспечивают реализацию приобретаемого в обучении
поведения [Edelman, 1989] (см. также гл. 14).
В силу ряда обстоятельств (таких, как доминирование рефлекторных представлений о
работе мозга, технические сложности длительной регистрации нейрональной активности у
свободно подвижных животных и др.), основной экспериментальный материал для
построения психофизиологической теории научения был получен в экспериментах, в
которых научение не было непосредственным предметом изучения. Так, на основе
исследования способности животных к экстраполяции Л.В. Крушинский предположил
наличие резерва нейронов, обеспечивающих фиксацию нового опыта [Крушинский, 1977].
В экспериментах, исследовавших нейрональные основы зрительного восприятия, было
обнаружено существование нейронов, специфически активировавшихся при
предъявлении конкретных лиц или определенных черт лица. Дополнительный анализ,
проведенный авторами, позволил утверждать, что эти клетки отвечают представлению о
гностических нейронах [Perrett et al., 1982]. Этот термин был гипотетически введен Ю.
Конорски в его теории инструментального обучения для обозначения клеток, отвечающих
«отдельным восприятиям» [Конорски, 1970] и фиксирующих элементы опыта. Нейроны,
специфически активировавшиеся при предъявлении конкретных слов, были обнаружены в
экспериментах на человеке [Heit et al., 1988].
В других экспериментах, направленных на изучение нейронных механизмов поведения,
были обнаружены нейроны, специфически активировавшиеся при осуществлении
отдельных поведенческих актов [Ranck, 1975] или нахождении животного в определенном
месте экспериментальной клетки [O’ Keefe, 1979]. Последние были названы нейронами
«места», и на основе регистрации активности таких клеток у крыс при обследовании
новой территории было показано, что они рекруитируются из нейронов, «молчавших» до
помещения животного в новую ситуацию [Wilson, McNaughton, 1993]. Утверждение о
специализации «молчащих» нейронов в процессе формирования инструментального
пищедобывательного поведения кроликов было также высказано на основе сравнения
наборов поведенческих специализаций нейронов до и после формирования новых
поведенческих актов [Горкин, 1987]. В этой работе, на основе сравнения среднего
количества активных нейронов при одном прохождении микроэлектрода через всю толщу
лимбической коры кролика во время выполнения пищедобывательного поведения до и
после доучивания, было показано увеличение количества активных нейронов после
обучения. Эти результаты подтвердили высказанное Л.В. Крушинским и позднее В.Б.
Швырковым [Shvyrkov, 1986] предположение о наличии резерва клеток,
обеспечивающего усвоение нового опыта и явились свидетельством в пользу селективных
теорий научения.
5. СИСТЕМНАЯ ПСИХОФИЗИОЛОГИЯ НАУЧЕНИЯ.
ПРОБЛЕМА ЭЛЕМЕНТОВ ИНДИВИДУАЛЬНОГО
ОПЫТА
В силу того, что, как уже отмечалось ранее, психическое связано с системными
процессами организации активности целого мозга [Швырков, 1978], появление в
репертуаре индивида нового поведенческого акта и соответствующего ему психического
состояния связано с реорганизацией всей мозговой активности. В то же время многие
теории, объясняющие реорганизацию мозговой активности в процессе научения,
оперируют с локальными нейронными ансамблями, ограниченными одной или
несколькими структурами головного мозга [Eccles, 1977], и по этому параметру являются
скорее физиологическими, чем психофизиологическими.
На наш взгляд, наилучшим образом соответствует критериям психофизиологической
теории научения подход с позиции на основе теории функциональных систем,
предложенной П.К. Анохиным [Анохин, 1968], которая развита в работах В.Б. Швыркова
и его коллег. В соответствии с теорией функциональных систем (подробнее см. гл. 14)
любой поведенческий акт реализуется системой кооперативно действующих элементов
организма разной морфологической принадлежности, организуемой моделью будущего
соотношения организма и среды (результата поведенческого акта). Появление такой
функциональной системы в опыте индивида и соответствующего поведенческого акта в
его поведенческом репертуаре являются следствием научения и происходят в результате
процессов системогенеза, имеющих место как на ранних этапах онтогенеза, так и у
взрослого (см. гл. 14).
Подход с позиции теории функциональных систем позволяет по-новому поставить и
решить проблему единицы индивидуального опыта, понимая под ней некоторое
хранящееся в памяти и воспроизводимое целостное состояние субъекта, которая
приобретается в результате единичного акта научения. Дело в том, что эта проблема
привлекала внимание исследователей только на заре исследований, когда методы
экспериментирования ограничивались наблюдением и поведенческими экспериментами.
Позже все силы были направлены на описание процесса научения, а проблема элементов,
или единиц, субъективного опыта была отдана на откуп интуитивным представлениям
конкретного ученого. В тех же исследованиях, которые были посвящены строению
памяти и не могли обойти данную проблему, в качестве элемента обычно выступали
единичные стимулы среды и конкретные движения [Rolls, 1987], а для человека – еще и
семантические единицы [Эксперим. психол., 1963]. Из таких элементов строилась
гипотетическая матрица связей (ассоциаций), которая и должна была объяснить
извлечение из памяти и использование конкретного опыта в определенной ситуации. На
наш взгляд, слабым местом этих элементов с позиции психофизиологии является их
умозрительное выделение внешним наблюдателем и несоответствие целостным
пространственно-временным организациям активности мозга, которые только и могут
быть соотнесены с состояниями субъекта.
В то же время функциональная система поведенческого акта, реализующаяся
одновременно с множеством систем, которые сформированы на предыдущих этапах онтои
филогенеза, является конкретной формой системных процессов организации активности
мозга, и поэтому может быть сопоставлена с состоянием субъекта как единицей
субъективного опыта. Одновременно она является внутренним эквивалентом
поведенческого акта, который выступает в качестве единицы реального поведения. В
связи с этим вполне оправданным кажется предположение о том, что функциональная
система поведенческого акта является элементом субъективного опыта [Швырков, 1995].
В качестве одного из центральных положений теории функциональных систем является
представление о существовании множества уровней функциональных систем. Даже
биохимическую организацию сокращения отдельного мышечного волокна П.К. Анохин
рассматривал как функциональную систему. По-видимому, такая система не является
элементом субъективного опыта. Это противоречие было разрешено В.Б. Швырковым с
системно-эволюционных позиций через рассмотрение филогенетической истории
организма. Те функциональные системы, которые выступают в качестве подчиненных по
отношению к являющейся элементом субъективного опыта функциональной системе
поведенческого акта, были названы им прасистемами [Швырков, 1995], что обозначает их
соответствие целостным системам поведенческих актов, но у филогенетических предков
рассматриваемого индивида. Так, к примеру, будучи автоматизированным у человека, акт
дыхания выступает в качестве целостного поведенческого акта у моллюска.
6. ФИКСАЦИЯ ЭТАПОВ ОБУЧЕНИЯ В ВИДЕ
ЭЛЕМЕНТОВ ОПЫТА
Обозначив принципиальный подход к выделению элементов субъективного опыта, теория
функциональных систем легла в основу экспериментов по изучению субъективного
дробления поведения и исследования нейрофизиологических проявлений приобретения
нового опыта. Необходимость такого рода исследований была связана с присущей
любому исследователю произвольностью выделения отдельных актов поведения при
внешнем наблюдении. В этой ситуации в силу непрерывности поведенческого
континуума определение моментов смены поведенческих актов и, соответственно,
достижения их результатов полностью зависит от представлений экспериментатора об
организации поведения. Для устранения этого субъективизма было необходимо
исследовать организацию активности мозга в поведении и выделить специфические
параметры этой активности, соответствующие этапам смены и реализации
функциональных систем.
В связи с имевшимися в литературе данными о специфической связи активности
отдельных нейронов с движениями, целью действия, местом и другими характеристиками
поведения, в лаборатории В. Б, Швыркова была разработана экспериментальная модель,
позволявшая дифференцировать связь нейрональной активности с каждым из этих
аспектов поведения. Это была модель пищедобывательного поведения у кроликов,
которым для получения пищи из двух кормушек, расположенных в углах передней стенки
квадратной клетки, требовалось нажимать на педали, которые были расположены в углах
задней стенки той же клетки. Регистрация активности корковых нейронов кроликов в
сериях пищедобывательных актов вдоль каждой из боковых сторон клетки позволила
выявить специфические активации нейронов, сопровождавшие реализацию отдельных
актов данного поведения. Такие активации наблюдались во всех случаях реализации
соответствующего поведенческого акта. На основании этих результатов был сделан вывод
о специализации нейронов относительно определенных поведенческих актов, который лег
в основу концепции системо-специфичности нейронов [Швырков, 1995], утверждающей
принадлежность любого нейрона только к одной функциональной системе (см. гл. 14).
Наряду со специализациями нейронов относительно отдельных систем поведенческих
актов, таких, как «подход к педали», «захват пищи», «нажатие педали», также были
обнаружены и другие специализации. Во-первых, это специализации нейронов
относительно конкретных движений животного и, во-вторых – специализации
относительно объединений актов, например, подход и нажатие педали. Если первые
специализации можно было отнести к реализации прасистем, то вторые вступали в
противоречие с принципом системе-специфичности и, на первый взгляд,
свидетельствовали о наличии в субъективном опыте животного элементов,
объединяющих несколько систем поведенческих актов.
Это противоречие было разрешено в серии экспериментов с тщательным контролем
процедуры обучения. В этих экспериментах проводилось контролируемое поэтапное
формирование пищедобывательного цикла [Горкин, 1987]. Каждый из пяти этапов
обучения (обучение захвату пищи в конкретной кормушке; повороту головы в сторону
педали; подходу к педали; нахождению в углу педали и, наконец, ее нажатию) проводился
в отдельный день и состоял из 100–200 подкрепляемых подачей пищи реализаций
соответствующего этапа. Последующая регистрация активности корковых нейронов в
дефинитивном поведении позволила выявить клетки, специализировавшиеся на
отдельных этапах обучения.
Для всех этапов были обнаружены группы специализированных нейронов, специализаций
же, не соответствовавших этапам обучения, за исключением нейронов прасистем,
выявлено не было. При этом специализации относительно этапов, служивших в качестве
промежуточных поведенческих актов и отсутствовавших в дефинитивном поведении,
выражались в виде активации, перекрывающих два последовательных поведенческих
акта. Так, например, специализация относительно этапа нахождения в углу педали
проявлялась в активации, начинавшейся в конце акта подхода к педали и продолжавшейся
во время ее нажатия. Эти наблюдения позволили сформулировать вывод о специализации
нейронов относительно этапов обучения.
Этот вывод, по мнению автора, исключительно важен для объективного изучения
структуры субъективного опыта, так как дает ключ к выявлению элементов опыта,
выделяемых его обладателем. В то же время полный набор поведенческих специализаций
нейронов соответствует субъективному репертуару поведений. Таким образом, можно
заключить, что с точки зрения системной психофизиологии элементом индивидуального
опыта является функциональная система поведенческого акта, сформированная на
конкретном этапе научения, т.е. в опыте индивида зафиксирована история его
приобретения.
Как уже отмечалось, элементы одной функциональной системы поведенческого акта
расположены в разных структурах мозга и достижение результата происходит за счет их
согласованной, кооперативной активности. Однако количество элементов,
принадлежащих одной функциональной системе, в разных структурах и представленность
разных систем в виде соответствующих специализаций в конкретной структуре мозга
оставались предметом изучения. В связи с этим необходимо рассмотреть участие
нейронов разных структур мозга в формировании новых функциональных систем.
В экспериментах на уже упоминавшейся модели пищедобывательного поведения
кроликов было проведено сравнение наборов специализаций в различных областях коры в
дефинитивном поведении, а также сопоставлены изменения этого набора в моторной и
лимбической областях коры в результате доучивания животного. Проведенное
исследование показало, что хотя нейроны одной специализации могут быть обнаружены в
разных областях, их количественная представленность сильно варьирует. Максимальное
количество нейронов, специализированных относительно актов сформированного
обучением в экспериментальной клетке поведения, было обнаружено в лимбической коре
(около 40% зарегистрированных нейронов), в то время как в других корковых областях
они были либо единичными, либо не достигали и 10% [Шевченко и др., 1986].
Сравнение наборов специализаций до и после доучивания также показало существенно
большие изменения в лимбической коре по сравнению с моторной [Горкин, 1987]. В
лимбической коре после доучивания появились новые специализации, соответствовавшие
актам добавленного поведения. Специально проведенная серия экспериментов с
подсчетом количества активных нейронов в лимбической коре до и после доучивания
показала абсолютное увеличение их количества в результате доучивания. Эти данные
были восприняты как свидетельство специализации в обучении «молчащих», т.е. не
дающих спайковых разрядов, нейронов. При этом количество клеток,
специализированных относительно наблюдавшихся до доучивания актов, не изменилось.
Наоборот, в моторной коре появились лишь единичные новые специализации, зато
количество активных клеток прасистем достоверно изменилось.
Полученные результаты соответствуют представлению о перестройке опыта в результате
включения в его состав новых элементов. На нейрофизиологическом уровне анализа
результатом научения является формирование «добавки» к набору специализированных
нейронов и изменение использования прасистем при реализации прежде существовавших
функциональных систем.
7. ВЛИЯНИЕ ИСТОРИИ НАУЧЕНИЯ НА СТРУКТУРУ
ОПЫТА И ОРГАНИЗАЦИЮ МОЗГОВОЙ
АКТИВНОСТИ
Субъективный континуум, как и континуум поведения, представляет собой линейную
последовательность сменяющих друг друга состояний, которые соответствуют актам
поведения. Эти смены одних наборов функциональных систем на другие связаны с
достижением результатов одних актов и возможностью реализации следующих актов
поведения. В силу того, что субъект выступает в качестве активного компонента
соотношений организма со средой (см. гл. 14), переход к реализации следующей в
последовательности функциональной системы происходит за счет ее извлечения из
памяти под воздействием внутренних факторов. Факторы, определяющие разные
состояния систем и смену реализуемых систем в континууме поведения, были названы
межсистемными отношениями.
В соответствии с включенностью конкретного акта в жестко фиксированную
последовательность, такую, как, например, врожденные стереотипные формы поведения
или аппетентная стадия пищедобывательного цикла, эти отношения могут иметь жестко
детерминированный характер. В то же время (как в случае поисковой активности), эти
отношения не детерминированы и выбор происходит среди слабо связанных в памяти
функциональных систем. Но и жесткая детерминация отношений двух последовательных
актов внутри цикла не является однозначной – всегда имеется возможность альтернативы,
т.е. перехода к каким-либо другим актам поведения. Подобная возможность реализуется,
например, при насыщении и переходе от пищедобывательного поведения к комфортному
(отдыху) либо при переходе к ориентировочному поведению в связи с изменениями в
среде.
Можно утверждать, что смена актов происходит в результате выбора из ряда альтернатив
с разной вероятностью в поведении. Наблюдение с этих позиций за формированием
умения играть в многоклеточные «крестики-нолики» позволило создать модель структуры
памяти и межсистемных отношений, хорошо описывающую выбор хода в конкретной
игре [Александров и др., 1997].
Исследование нейрональной активности в поведении показало, что, наряду с ярко
выраженной специфической активацией специализированных нейронов в специфическом
поведенческом акте, в их активности имеется дополнительная структура. Эта
структурированность неспецифической активности (т.е. активности при осуществлении
других актов поведения) проявляется в наличии слабых активации и тормозных пауз,
сопровождающих реализацию других актов поведения. Связь таких изменений с
конкретным актом имеет, как правило, вероятностный, но достаточно устойчивый (по
статистическим критериям) характер.
Исходя из принципа системе-специфичности нейрона и его специализации относительно
элемента индивидуального опыта, было выдвинуто положение о разных состояниях, или
степенях актуализации элемента опыта в реальном континууме поведения.
Нейрофизиологическим выражением степени актуализации специфической
функциональной системы при подобном подходе является текущая частота активности
специализированного нейрона, варьирующая от нулевых значений при исключении
системы из состояния субъекта поведения до максимума при реализации
соответствующего акта поведения.
Таким образом, на основе анализа нейрональной активности был сделан вывод об
актуализации множества функциональных систем при осуществлении конкретного акта
поведения. Обнаружение определенной структуры актуализации конкретной
функциональной системы в поведении позволило использовать регистрацию активности
нейронов известной специализации для изучения отношений конкретного элемента опыта
с другими элементами.
В качестве метода выявления межсистемных отношений используется построение
паттернов активности специализированных нейронов во всех актах анализируемого
поведения и распределения вероятности активации нейрона в конкретном акте поведения.
За активацию принимается не менее чем 1,5-кратное превышение «фоновой» частоты
активности. Эти формы представления нейрональной активности иллюстрируются на рис.
15.1 в сравнении с распространенным методом представления активности в виде растров.
Особенно яркие результаты дало использование данного метода для решения проблемы
отражения истории научения в организации межсистемных отношений.
В силу фиксации этапов научения в виде элементов субъективного опыта история
научения индивида является содержанием его опыта. Однако кроме состава элементов
опыта, история его формирования может (наряду с логикой поведения) определять
отношения систем.
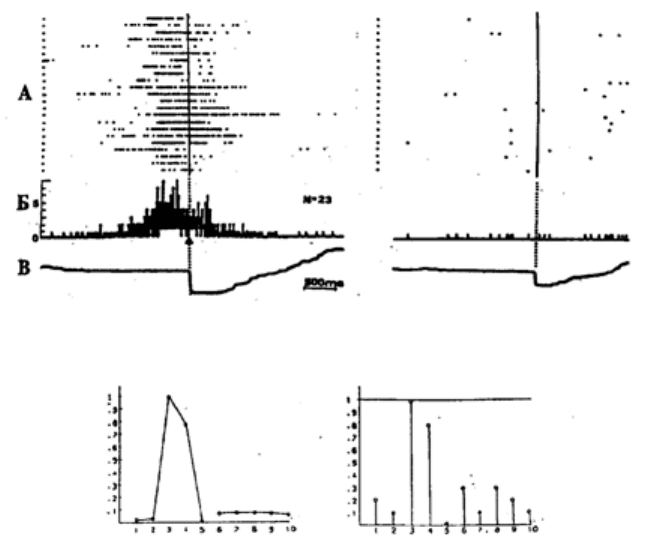
Рис. 15.1. Разные способы представления активности нейрона
А – растры активности нейрона лимбической коры; Б – гистограмма; В – усредненная
отметка нажатия педали (слева–левой, справа–правой), относительно начала которой
совмещены растры и построены гистограммы. Внизу: графическое представление
активности того же нейрона. На левом графике – картина распределения средних частот
(паттерн) активности нейрона в актах циклического поведения. По оси абсцисс – номера
актов, по оси ординат – нормированная средняя частота активности. На правом графике –
вероятности наличия активации в тех же 10 актах. По оси абсцисс – номера актов, по оси
ординат – вероятность активации нейрона в соответствующем акте
Было проведено специальное исследование активности нейронов лимбической коры
кроликов в пищедобывательном поведении, сформированном с применением разных
стратегий обучения. На рис. 15.2 схематически представлены использовавшиеся разные
стратегии обучения внешне одному и тому же циклическому пишедобывательному
поведению. Разница между стратегиями состояла в последовательности формирования
отдельных блоков поведения – поведения у кормушки и поведения у педали – на двух
сторонах экспериментальной клетки. Формирование отдельных поведенческих актов
производилось так же, как было изложено ранее. На рисунке последовательности этапов
научения показаны цепочками стрелок.
Для выявления сформированных межсистемных отношений была зарегистрирована
активность нейронов лимбической коры в дефинитивном поведении обученных таким
образом животных. Из всей совокупности зарегистрированных нейронов для анализа
были отобраны клетки, специализированные относительно сформированных обучением в
экспериментальной клетке поведенческих актов.